Embryology Biology 441 Spring 2012 Albert Harris
Cancer, from an Embryological Point of View
Streptomycin selectively damages bacterial ribosomes, and does much less harm to animal cell ribosomes.
To cure cancer, what is most needed is some chemical (or other treatment) that kills cancer cells much more than it hurts normal cells.
Almost no progress is being made toward that goal.
25% of the students in this course will get a fatal form of cancer; 5% will be cured by surgery and/or radiation and/or chemotherapy. 20% of you will die of the cancer...unless some revolutionary new direction of research occurs. But there is no indication of such a revolution. Nearly all cancer researchers are satisfied with things as they are.
They seek drugs that you must keep taking, over and over, to prolong your life, instead of a real cure.
They are satisfied to convert cancer to a chronic, life-long disease.
Drugs like Gleevec are designed to reduce the abnormalities of cancer cells
Prices are very high; for example see http://www.healthwarehouse.com/gleevec-400mg-tablets.html
Gleevec 400mg tablets, price for 30 tablets $5,736.50 !
Do the math! 5,736.50 / 30. That's a hundred and ninety one dollars per pill that costs maybe a dollar to synthesize, and was NOT invented by the drug company, and will not cure.
Gleevec works by inactivating an over-active enzyme that covalently binds phosphate groups to proteins.
Why not look for a drug that selectively kills any cells in which abnormal numbers of phosphates are bound to proteins?
Because then you wouldn't need to keep taking the pills!
Economic motivations are wonderful; they are the foundation of our freedom and national greatness; and they really, really work! If companies can make more money doing X than doing Y, then which will they do?
Suppose that X is finding a drug that cancer patients have to keep taking, over a long time, and won't cure them. Suppose that Y is finding an actual cure.
Will they really choose to do X and avoid doing Y? Will the public notice?
Abnormalities that have been discovered to occur in many (or all?) varieties of cancer cells (differences between cancer cells and the normal cells they are derived from).
To cure cancer, just invent a method that will selectively kill just those cells that have one or more of these abnormalities. You might very well succeed. Almost nobody has tried. They keep busy doing something else, which will be described later.
1) Glucose uptake is much higher than normal in most cancer cells. Related to this is an abnormally (very) large secretion of lactic acid, and abnormally low amounts of ATP production by mitochondria. This set of inter-related abnormalities was discovered around 1930 by Warburg, who had won the Nobel Prize for previous discoveries, and is one of two (unrelated) phenomena both called "The Warburg Effect". Excess glucose uptake into cancer cells is the reason why PET scans can detect cancer, and reliably distinguish cancer cells from normal cells. I have had several PET scans myself, and they accurately mapped the tumors. They cost about three thousand dollars per scan.
The biochemical cause of these phenomena remains unknown, and very little research has been done on it, because it doesn't fit in with oncogene research. The best English language textbook on cancer biology doesn't even mention the Warburg effect. Almost no research has been done about how to kill cancer cells based on either their anaerobic metabolism, or the excess uptake of glucose. (Despite very large amounts of research on how to use the phenomenon to map tumors).
2) MRI images of cancer cells are dramatically different than normal tissues. This was the subject of the first research paper about MRI. Nobody ever discovered the reason why cancer cells' MRI images are abnormal. Almost no research has been done on the subject.
3) The acto-myosin cytoskeleton of cancer cells is disorganized, and they exert much weaker traction forces as they crawl.
4) Cancer cells can crawl onto less adhesive substrata, from more adhesive substrata. This is a loss of haptotaxis. It was discovered by Prof. Barbara Danowski, as part of her PhD research in this department. She invented the experiment herself. She then did important research at the University of Pennsylvania, and is now Professor and Chair of Biology at Union College in New York. I had never considered the possibility, until she proved it.
5) Defective cell-cycle checkpoint controls. For example, cancer cells tend to go ahead and copy DNA, even if it is damaged. Normal cells would delay starting to copy their DNA until damage to it has been repaired.
6) Excessive inhibition of apoptosis. In particular, 95% of cases of the "follicular" kind of Non-Hodgkin's lymphoma are caused by a breakage and incorrect re-joining of the 14th chromosome and the 18th chromosome, in such a way that the promoter region of the antibody heavy chain gene is next to a gene named bcl-2. This stands for b-cell lymphoma two, because it was the second lymphoma-causing gene to be discovered by looking for chromosome translocations in cancerous lymphocytes of human patients. Nematodes have a gene almost identical to human bcl-2. It is needed to inhibit programmed cell death, both in vertebrates and in nematodes (and probably all animals). Experimenters have even spliced the human DNA sequence for bcl-2 into nematodes, in place of the nematode's own gene. The human gene works fine in nematode embryonic development.
When a human b-lymphocyte has this particular chromosome translocation, it makes bcl-2 protein instead of antibody, and can't undergo programmed cell death. Although such lymphocytes don't grow or divide any faster than normal lymphocytes, they do accumulate without limit. Eventually, they displace your normal lymphocytes and fill up your bone marrow, until you become anemic, can't make antibodies etc. and die.
Ironically, the treatment is often a chemical designed to kill fast-growing cells by damaging their DNA. It was assumed the cancerous lymphocytes must be growing too fast, and that cross-linking their DNA would selectively kill the fastest growing lymphocytes.(That, itself, isn't very logical. Why should it kill a cell to prevent it from doing something abnormal? If they grew slower than normal, would you expect to be able to kill them by speeding them up?)
Consider the following hypothesis about how chemotherapy really works: When DNA is damaged by radiation or a drug like cyclophosphamide, non-cancerous cells detect the damage and slow down their growth, halting at a check-point until the damaged DNA has been fixed. The cancerous cells, however, because their check-point controls are broken, continue to copy DNA and undergo mitosis. This failure to stop is really what kills them. This is sort of the opposite of what had been assumed. Cancer cells die because they can't stop, not because they grow too fast. Any time a drug works, there must be some reason; but the true reason may be very different than what the drug was designed to do.
7) Lack of anchorage dependence, the ability of cells to survive and continue dividing without being spread out on a solid substratum. Cancer cells can grow and survive suspended in a gel. Non-cancerous cells will undergo programmed cell death if you culture them on an area of substratum that is smaller that the area they normally occupy when attached to a Petri dish. Nicholas Maroudas and Donald Ingber have done the best research on this.
8) Increased secretion of proteolytic enzymes (metalloproteases, etc.)
9) Loss of differentiated characteristics ("tumor progression")
10) Gain of abnormal combinations of gene transcription
NOT excessive rates of cell growth! That is a myth that has held back cancer research, worse than anything else. It's boloney to claim that "cancer is caused by cells growing too fast; and chemotherapy works by slowing down this growth". But there are dozens of web sites that claim this stupid error.
There are more kinds in addition; nobody even knows how many more there are, and almost no research or funding goes toward searching for more.
----------
I) The disease cancer results from a cell of your own body changing so that it and its mitotic daughter cells divide and crawl without limit.
It is normal for body cells to grow and divide, and to crawl from place to place.
But cell growth and cell locomotion are normally controlled, by embryological mechanisms, which are only partly understood.
"Contact inhibition" means the normal inhibition of cell growth by crowding, and directional inhibition of cell crawling where once cell touches another.
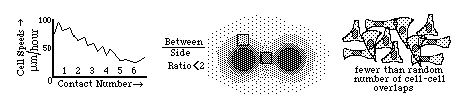
Diagrams of the quantitative experimental tissue culture measurements done by Michael Abercrombie and Joan Heaysman, by which they discovered and proved the existence of contact inhibition of cell locomotion (crawling). By these criteria, they also discovered that cancer cells in tissue culture have less sensitivity to contact inhibition; that seems likely to be at least part of the reason for their increased invasiveness. Other researchers later used the phrase "contact inhibition" to mean reduced rates of cell growth and mitosis in crowded tissue cultures. Cancer cells are likewise less inhibited than equivalent normal cells.
For many years, other scientists confused these two uses of the phrase "contact inhibition" (inhibition of growth as well as locomotion), assumed the same mechanism caused both, and that cancer cells always lacked both (which Abercrombie never claimed). Then people over-reacted in the opposite direction. The inhibition of locomotion now seems to be caused by prevention of actin fiber assembly near where cells touch each other. Much more research is needed to find out whether cancer cells are more able to continue crawling where they touch other cells, whether this is related to their uncontrolled growth, or if such abnormalities can be targeted by new kinds of chemotherapy.
These mechanisms of inhibition of growth and of locomotion may or may not have the same (or overlapping) mechanisms.
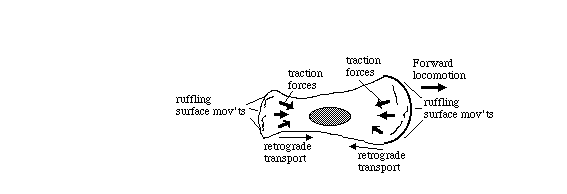
Tissue cells (including body cells in tissue culture) crawl by means
of traction forces exerted behind their leading edges. Traction
forces are exerted mostly behind areas where the plasma membrane
bends, bubbles and fold irregularly in a process called "ruffling"
(as can be seen in the video time lapse sequences below).
These cell surface movements were was once thought to be peristaltic, but
now are now known to be caused by continual re-assembly of cytoplasmic actin.
Part of contact inhibition is the paralysis of ruffling where two cells touch each other,
as shown in the diagram below.
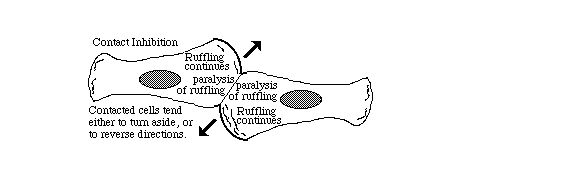
Many tissue cells behave as if actin assembly is somehow turned off where one cell touches another. Such cells behave as if the mechanism of locomotion is inhibited next to cell-cell adhesions. Sometimes one or both cells turns or crawls away from the cell-cell contant; and sometimes new cell-cell adhesions form, and the adhering parts of both cell margins cease locomotion and ruffling.
II) The great majority (>95%) of human cancers are caused by somatic mutations in a few specific genes (called "Oncogenes").
In many other species, large fractions of cancers are caused by communicable viruses. (examples include cats and turkeys)
This department gives a very good course specifically about oncogenes and how they cause cancer.
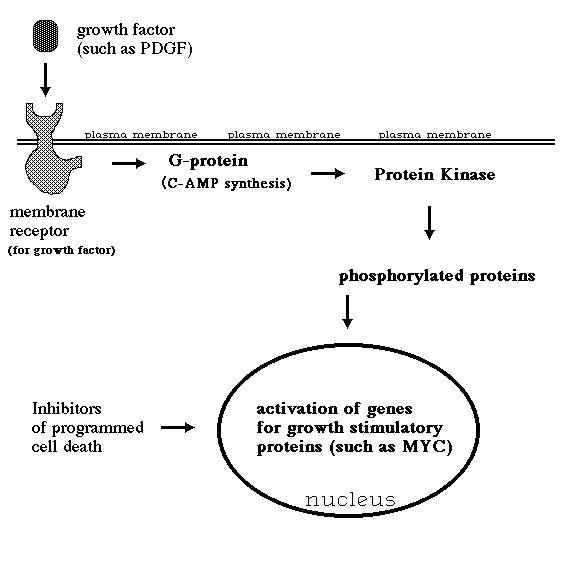
III) Several sexually transmitted papilloma viruses cause cervical cancer in women. A vaccine has recently been developed which inhibits infection by these cancer-causing papilloma viruses.
Many newspapers etc. have confused this with 'a vaccine against cancer.'
An editorial in the News and Observer "explained" that the vaccine "Is not yet approved for boys." but doesn't ask "Why not?".
These viruses infect both sexes: but the cancers they cause are in females.
What is their reasoning for vaccinating only one sex? After all, from whom do females catch these viruses?
IV) Cancer cells retain many of the properties of whichever differentiated cell type they began from:
