The "Embryology" of Plants
The mechanisms by which plants form their bodies make an interesting contrast to the mechanisms used by animals. We can understand more about both animal and plant development by contrasting the two. In particular, notice the tremendous economic (and sometimes military) power that was gained by discovering the specific chemicals that plants use as signals to control growth and differentiation. This occurred long before current molecular genetic methods. What will happen if people discover methods for exerting the same degree of control over development of the human body, as was achieved by the discoveries of plant "hormones" such as auxin, in the 1930s-60s.
Similarities of plant development to that of animals:
-
* Plants have many differentiated cell types (but more often can switch from one to another)
* There are consistent embryonic stages (would you believe there is a "torpedo stage"!?)
* Asymmetries sometimes depend on external signals, and in other species are created in the ovary.
* There is such a thing as plant tissue culture.
* You can make chimeras, and even fate maps.
* There are homeotic mutants!
The most-used "model organism" for genetic analysis is Arabidopsis thaliana, "The Wall Cress" fast life cycle, accumulation of many mutant strains & very small genome (totally sequenced).
If you prefer one-celled plants, Chlamydomonas is the best model organism. It has also been sequenced, has many mutants that alter flagella, and is almost unique among plants for being able to survive mutations that prevent photosynthesis. Do you see why that is important?
Differences between development of plants, as compared with animal embryology:
* NO cell locomotion, no invagination, or any other morphogenetic cell movements.
* Cell walls made of cellulose, lignin & other stiff materials are secreted around all cells.
* Cells can grow larger by osmotic swelling; and this swelling can be much more in some directions than in other directions, producing changes in shape, often major changes in shape.
* Morphogenesis is produced by controlled weakening of cell walls, in certain places and directions. and by the spatial control of differentiation; & note that differentiated cells stay put!
* Dedifferentiation can allow cells to change cell types; cuttings can form whole plants
* Cell division occurs by secretion of a new cell wall across the middle of the cell, instead of by contraction of an actin & myosin ring. But actin & microtubules are involved, somehow.
* Non-determinate growth. Instead of growing to a certain shape and size, plants just keep on growing, and develop different individual shapes, often strongly depending on the environment. Although that makes them a lot different from us and all other vertebrates, it is really not so fundamental as people tend to think. It has to do with being non-motile. Notice that corals, sea fans, bryozoans, sea squirts, sponges and many other vertebrates also keep on growing just like plants. And sea squirts are members of the same phylum as us: Chordata. If there were some kind of vertebrate that was sessile, its growth would probably also become indeterminate. What do you think?
* Seeds are (or contain) partly developed plant embryos; they are not equivalent to oocytes. Imagine that our embryos developed up to, say, the late cleavage stage, or maybe to the gastrula or neurula stage, and then stopped there for a long time in a dormant state of inactivity. That would be analogous to seeds, which are highly multicellular, and part-way along in their embryonic development.
In fact, there is a special kind of Australian fresh-water fish whose embryos "hibernate" at about gastrula stage (in case you are interested)
A peanut is a partly developed seedling, with the two big halves each being a cotyledon (= special embryonic leaf that stores food, & are first two leaves the baby plant produces) That tiny thing at one end between the halves of a peanut is the plant embryo itself, partly developed.
Plant embryos develop part way, and then wait in a state of dormancy until conditions are right!
Mitoses are concentrated almost entirely to certain regions called meristems.
A shoot meristem at the top end of each growing shoot, and each branch.
Aroot meristem at the bottom end of each root and root branch.
And lateral meristems around the sides (in larger plants); a lateral meristem is called a cambium. one cambium produces xylem from its inner face and phloem from the outer side. and bark is produced by a cork cambium.
The cells of meristems tend to be small, with small vacuoles;
But as cells differentiate, they usually expand greatly, and develop huge vacuoles that take up a large percentage of the cell volume.
A latent inactive potential (non-growing) meristem forms at the locations just above where each leaf is formed. These are inhibited from becoming new shoot meristems by auxin, one of the plant hormones.
Auxin and other plant hormones:
Plant development is controlled by gradients of specific chemicals called plant hormones. Please notice that these are mostly rather different from animal hormones, because their concentrations are often kept very different from one part of the plant body to another. Both plants and animals use hormones for long range signalling, but plants use some of them to control exactly where certain structural changes will occur. If animals were found to have chemicals that could, for example, cause a new leg to grow as a result of making this chemical more concentrated in a certain place, then embryologists would probably cause that chemical a "morphogen", rather than a hormone.
Plant hormones have small molecules with relatively simple molecular structures (& water soluble). (You DO NOT have to memorize any of these structures however!)
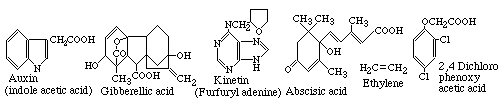
You should learn the names of the five major kinds of plant hormones:
Auxin, Ethylene, Kinitin, Gibberellic acid, and Abscisic Acid.
The chemical structure of one of the artificial chemical analogs of auxin (2-4 D) is also drawn above; plants never make this, but they do respond to it. This is an example of one of the most important practical facts about biochemistry, which is that if you make artificial chemicals that have approximately the same shapes and charge distributions as some chemical that an organism uses for an important function, then this artificial chemical is likely to "gum up the works" by binding to the receptors (or enzymes, etc.) meant to bind the normal compound. Most of pharmacology is based on this principle; and so are most herbicides.
These chemicals were discovered long ago by basic research.
Their use is valued at billions of dollars per year: for making plants grow, die, ripen etc. when and where humans want them to.
If equivalent chemicals for controlling animal development are discovered: what will the uses be?
Several other plant hormones have also been discovered more recently. These include brassinosteroids (which are actually steroids, with similar chemical structures to certain well-known animal hormones). Another plant hormone is jasmonic acid, and still another is a peptide called systemin. I thank my colleague Alan Jones for telling me about these other hormones. He had made important discoveries about receptors for plant hormones. He is one of several excellent researchers on plant development in this department. If you like this subject, you might consider an honors project with one of them.
Plant shoots do the following 5 things: all of which turn out to be controlled (mostly) by auxin
a) Bend toward light. (positive phototaxis);
b) Elongate more in darkness than in light
c) Bend upward relative to gravity (negative geotaxis);
d) Inhibit branching near a growing tip.
e) Sprout roots at the bottom.
Auxin is made and secreted by the cells at the shoot tip. Auxin diffuses and is actively transported downward from there. (The cells somehow detect gravity, and pump auxin toward whichever side is down) If a plant is leaning sideways, this downward transport will cause the concentration of auxin to become larger on the lower side and smaller on the upper side. Auxin causes cell walls to weaken, so that osmotic pressure stretches them, and thereby causes cells to grow larger where auxin concentrations are larger. This makes leaning plants grow more on the lower side, therefore turning upward. Auxin is decreased in concentration by bright light, either by breakdown or by active transport away from the light. Plant stems therefore bend toward light. Auxin inhibits development of new shoot meristems (at those locations just above where leaves were formed); therefore new shoots are inhibited from sprouting near growing tips. Cutting off a shoot tip removes this source of auxin, thus de-inhibiting formation of new branches. Several new shoot meristems tend to form simultaneously, if you prune a hedge or other plant correctly. These had all been inhibited from growing by the auxin coming from the old tip. For similar reasons, political dictatorships often produce multiple co-dictators each time the old one dies.
Although auxin inhibits sprouting of shoots, it has exactly the opposite effect on roots, and can even convert other cell types to root cells. The downward transport of auxin is what causes cuttings to form roots at their bottom end. The chemicals sold in garden stores to promote rooting are made of auxin, or sometimes chemical analogs of auxin.
The discovery of auxin began with Charles Darwin, who did experiments on the ability of plants to bend toward light. By cutting off shoot tips, he found that bending toward light does not occur unless the tip is present. However, when he tried the effect of wrapping metal foil around different parts of growing plants, he discovered that the shoot tip does not seem to be the part that actually senses the light. Instead, it is the region below this which you need to shield from light in order to prevent the phototaxis phenomenon. Darwin hypothesized that some chemical is secreted at the tip, but then travels downward, and it then somehow altered or inactivated by light, and causes bending by some effect on cell growth in this lower region. I don't know whether he foresaw synthesizing the chemical, or making chemical analogs.
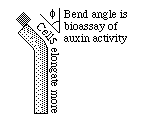
This chemical remained hypothetical from the 1870s until the 1930s & 40s. Its chemical structure was discovered by means of clever bioassays that measured small amounts of differences in the amount of cell elongation stimulated by different chemicals. At one stage of research, shoot tips were cut off and set on tiny blocks of agar gel; and these agar blocks were set on top of the cut-off shoots. Auxin was able to pass through the agar blocks and stimulate elongation of the plant tissue below them. When such agar blocks were set on one side of the cut-off tips, then there was more cell elongation on the side receiving the agar. Eventually, different chemicals extracted from plant tissues were fractionated (by techniques such as chromatography), and these chemicals were compared with one another to see which could produce the most bending by differential cell elongation. The chemical with the greatest effect turned out to be indole acetic acid. The name "auxin" had been invented many years before. This is often true in biology; the name was invented based on a theory about the mechanism; and a bioassay was invented to identify which chemical had the hypothetical effect. Serotonin (an animal hormone) also was named first, and then its chemical nature discovered by a simple bioassay using strips of muscle tissue. Many molecular biologists have very distorted ideas about mechanisms get discovered. The story of auxin is a good example of how discoveries often occur: indirect evidence, followed by hypotheses, followed by bioassays, following eventually by synthesis of chemical analogs. Prozac is a chemical analog of serotonin, like 2-4D is of auxin.
Question for discussion: Can you invent some other bioassays, that would have compared relative amounts of auxin in extracts based on some of its other effects, instead of being based on shoot elongation?
A supposed flower-inducing hormone, "florigen" was postulated & looked for by many scientists. But it probably does not exist!
Plants have an analogue to programmed cell death in the formation of abscission layers at the bases of leaves, fruits and flowers. The death of the cells in these layers allows the structure to fall off.
The strengthening of our muscles and bones in response to stress has a plant analogue in the formation of reaction wood. In conifers, this forms in response to compression; but in dicots in response to tension! It is very strong, although tending to warp; people used to make archery bows out of it.
The geometric arrangement of new leaves at a shoot tip, or of the petals of a flower is called phyllotaxis. For example, many plants form pairs of leaf buds directly opposite from one another, with each new pair 90 degrees from the previous pair. Most form leaf buds in a spiral pattern in which the angular displacement approximates 138 degrees. This story involves "golden ratios", "Fibonacci numbers", and too much mysticism. Surgical removal of early leaf buds can cause the next ones to form in different places than they otherwise would have; & local stimulation by implanting a hormone-impregnated bead can reverse the spiral direction.

Historically, observations of developing flowers seem to have played an important catalytic role in the development of early embryologists' thinking about such things as the primary germ layers (which were called "germ leaves" (= "Blätter") by the German embryologists who invented the germ layer concept) They were very impressed by the graded similarities between leaves, sepals and petals and other homologous structures in developing flowers and fruits. This analogy helped them to recognize the existence of the 3 germ layers in bird embryos, and helped people move beyond preformationism.
Some questions that you ought to be able to answer on an exam:
Name 5 examples of plant hormones.; How do these hormones differ from animal hormone, including differences in function, differences in molecular nature & differences in distribution in the body?
What effects can be produced by chemical analogs of these hormones? What is the common name for a widely-used chemical analog of auxin? How was auxin identified? What bioassay was used to distinguish which chemical fraction contained the active ingredient? Could you invent several other bioassays based on different effects of auxin that are now known? What did the chemical nature of auxin turn out to be? & how does it differ from being a purine, although the chemical structures are somewhat similar. Which of the plant hormones does have a purine structure? Are any of these plant hormones steroids? Are any of them peptides or proteins? (& what about hormones used by animals?) Are one of the plant hormones a gas? Why does one rotten apple spoil the whole barrel-full? Does it matter if gas can leak out of the barrel? What gas can be used to force fruits to ripen? What plant embryonic structure is kind of like the yolk sac of a bird or reptile embryo? What are some differences between plant development and animal embryology? Are some of these differences more fundamental that others? Is the differentiation of plant cells more or less reversible than in animals, as a rule. How is that difference related to the ability to "root" small cuttings taken from plants? Why does trimming a hedge make it sprout thicker? How might you prevent that thickening with a certain herbicide? How might you use low concentrations of that same herbicide to promote rooting of shoots? Why do new roots grow at the bottom of a cutting? Would that still happen in space? How do plants bend toward light? How is the mechanism of this bending related to how they grow very long and spindly in total darkness? What pigment do plants use to detect day length? When flowering is controlled by day length, is it really night length that is detected? How could you tell?
The earliest stages of development are very difficult to study in dicots & other higher plants;
mostly because these stages occur inside the ovary of the mother plant.
Instead, one can study the equivalent stages in the brown alga Fucus as a model system ("kelp")
They release oocytes directly into the sea water; sort of the plant kingdom's equivalent to sea urchins!

The 1-cell stage embryos develop their future rhizoid-apical polarity in response to any of many different stimuli, including light direction (rhizoid to dark), pH gradients (rhizoid toward acid), gravity (rhizoid down) and electric fields; Research in this department discovered that cytoplasmic actin transports material to future rhizoid (root) end, where it is added to cell the wall; Certain components of the cell wall in the rhizoid end then stimulate further rhizoid growth.
Volvox is used as a model system for an extremely primitive evolutionary stage of multicellularity
They amount to a colonial form of Chlamydomonas
The floral meristem develops into flowers: a flower is made up of 4 kinds of structures, all of which are homologous to leaves
sepals (= the leaf-like structures outside & below the petals)
petals (you know what they are)
stamens (the male sexual structure; a rod-like thing that produces pollen at its end)
carpals (the female sexual structures, usually 2 or more fused together; oocytes produced inside them)
Various homeotic mutations are known that result in tissues that would have developed into one of these forming a different one instead.
For example, in one of the more boring ones, leaves develop in place of sepals, petals, stamens & carpals
But there are some more interesting & tantalizing patterns seen in phenotypes of other homeotic mutants.
In agamous mutants, petals develop in place of stamens and sepals instead of carpals!
(instead of the normal sepal, petal, stamen, carpal, you get sepal, petal, petal, sepal.)
In apetela2 mutants stamens replace petals, and carpals replace sepals
(instead of the normal sepal, petal, stamen, carpal, you get carpal, stamen, stamen, carpal)
Perhaps the most interesting kind of homeotic mutations are those of the "cauliflower" kind, in which each sepal, petal etc. tries to develop into a whole flower, and then each sepal, petal etc. of those flowers ITSELF tries to develop into a whole flower. An analogy in humans would be if each of the 5 fingers of an embryonic hand tried to develop into a whole hand, and then each of the 25 fingers then developed as a separate hand, with all 125 of their fingers then trying to develop into a separate hand, and so on!
(like the self-calling algorithms that can be used in Pascal; has anybody read "Godel, Escher, Bach"?)
A somewhat analogous phenomenon is the formation of compound leaves.
There are mutations that increase the extent of compounding.
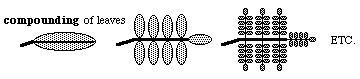
Plant developmental biology is an exciting field with room for new researchers; you might consider it!