Lecture notes for Monday, February 11, 2013
Tissues and organs that develop from mesoderm
| Lateral plate mesoderm |
Intermediate mesoderm |
Somites | Notochord | Somites | Intermediate mesoderm |
Lateral plate mesoderm |


|
We are members of the phylum "chordata", all of the members of which have a notochord at some stage of their development. In mammals and birds, the function of the notochord is replaced by the vertebral column (bones of the back-bone+intervertebral discs) But tadpoles and larval fish actually depend on the notochord to allow them to swim by alternating contraction of muscles along each side.
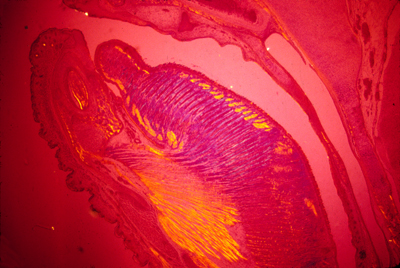
The notochord mesoderm also induces formation of the neural tube. The structure of the notochord is a long stack of (not always) flattened cells, that often have very much the geometry of a stack of coins. Wrapped tightly around the surface of this long cylinder of cells are layers of fibers of type I collagen in many spiraling layers, that alternate in direction (a clockwise layer, then a counterclockwise layer, then another clockwise layer, etc. etc. etc.) The cells inside the notochord then form large vacuoles in their cytoplasm, which makes each cell swell in volume, pushing against the spiral layers of (strong & non-stretchable!) collagen fibers. The result is that the notochord can bend side to side, but can't be compressed along its anterior-posterior axis.
Tail of a living tadpole. The broad line just below the center
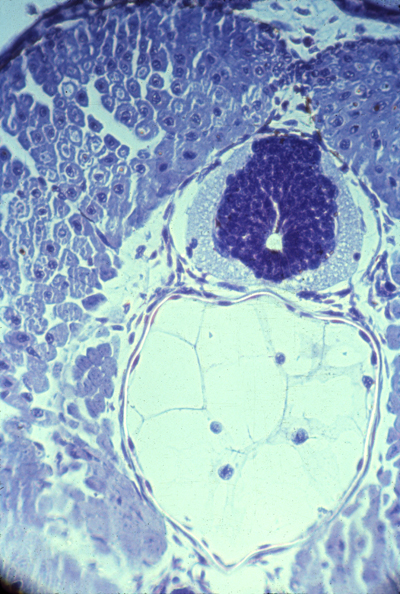
Neural tube and notochord
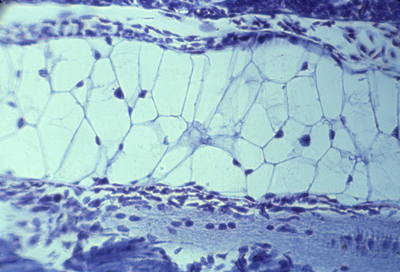
Notochord of salamander
Section through chick embryo
A short digression about the VERY important protein, collagen.Type I collagen is the main extracellular protein of the body, and causes the mechanical strength of tendons, skin, organ capsules, the wall of your eyeball (including even the cornea!).(even bones are 1/3 type I collagen).
Jello (flavored gelatin) is type I collagen; so is Elmer's glue.
Collagen can be purified from rat tail tendons, and skin of cows,
A series of very similar proteins are named "type II collagen"
type III collagen, type IV collagen, and guess what comes next?
For many years, it was an unsolved problem how type I collagen gets arranged into so many different geometric patterns in the body . In the early 1980s, a UNC grad student and I did experiments that (maybe?) gave the answer to this paradox. We said that mesenchymal cells pull strongly on collagen, and mechanically rearrange it according to what forces are exerted at each location.
Photograph of a rat collagen gel compressed by chicken cells
Collagen aligned in petri dish
Theory of tendon formation
Photograph of fluorescently-labeled rat collagen
When fluorescent collagen was injected into developing chicken embryos, then it got made into many different anatomical structures:
Photograph of chicken artery, with fluorescent rat collagen wall.
Next, back to questions about how the notochord forms.
The best research about this has been done by Ray Keller,
(Currently the chairman of the Biology dept at UVA) They describe a process of convergent extension, in which the future notochord cells extend processes sideways; these then adhere to each other, and these processes actively contract, pulling in the sides. It would have fit our theories a lot better if the collagen sheath were wrapped around the notochord as it first forms. Then cell traction could have tightened this collagen sheath, so as to constrict and elongate the notochord. But I have looked carefully at the scanning electron microscope pictures of the earliest stages of the notochord, and I have to admit that I don't see any sign of collagen fibers wrapping around it yet. A good experiment for somebody to do would be to inject fluorescently-labeled collagen into embryos in the area around the notochord at various stages of development, and observe whether (and at what stage) this collagen gets rearranged to form part of the notochord sheath. Please notice similarities between the fluorescent collagen wrapping around the chicken artery in the figure above as compared with the photograph earlier in the notes labeled "neural tube and notochord" where you can see a thin white line surrounding the vacuolated cells.Although this line wanders a little in the photo, I believe that's because of fixation and sectioning. In the living embryo, it should be stretched tight. Several odd facts about the notochord: In reptiles, birds and mammals, the notochord cells are the last to leave the surface (epiblast) at the Hensen's Node.
* In some species (humans & turtles), Hensen's node is partly an invagination, & a hollow tube runs down the middle of the notochord * In sections of amphibian embryos, you can see pigment granules tend to accumulate down the middle of the notochord. * The spiral layers of collagen fibers that are the sheath of the notochord are oriented at 90 degrees to each other. This is one of several examples in which collagen fibers mysteriously become oriented in alternating layers, with the fibers of each layer exactly perpendicular to the fibers on the layers above and below.
Also, there are many examples in which muscle cells become lined up in perpendicular alternating layers: Mammal tongues, elephant trunks,
squid tentacles, and in Hydra, both the body and the tentacles.
Polarization micrograph of a section through a mouse tongue
Anybody who could figure out the mechanism that creates these perpendicular layer patterns, and prove their theory true, would go down in biological history. Somites are segmental blocks that form beside the neural tube. At first, there are continuous columns of "paraxial" mesoderm
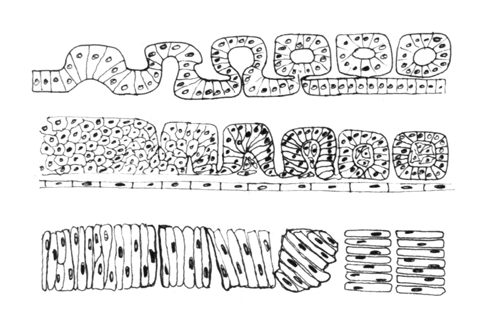
The drawing above shows the geometrical rearrangements that some scientists have reported to casue somite segmentation in different classes of chordates. Top: Amphioxus, Middle: bird and mammal, Bottom: frogs. Anterior is to right side.
Then these columns spontaneously split apart, one pair at a time.
Longitudinal section of notochord and somites in a developing frog embryo
Somites forming in a fixed whole mount chicken embryo. Anterior to left.
Somites forming in a living chicken embryo. Anterior to left.
The physical mechanism of splitting apart could be active constriction, or could be decreases in cell-cell adhesion proteins. In scanning EM photos, "somitomeres" can be seen, where a somite is going to form, apparently as the first stages of somite formation.
In most species, somites start as hollow epithelial balls,
and conversion of cells from being mesenchymal to epithelial
therefore seems to be part or the process of separation.
Much research has been done on the genes needed for somite formation The most popular categories of theory are 1) That somite segmentation has the same mechanism as formation of segments in flies, using genes analogous to "even-skipped" etc. 2) The "Clock-and-Wavefront Hypothesis", according to which one quantity oscillates higher and lower in amount, while another variable forms a gradient that gradually increases in amount along its length. A somite is supposed to be split off each time the oscillator increases. Really, no one even knows whether we should think of the splits as being the real entities, as opposed to the somites themselves.
For most species, the same number of somites forms in each embryo. You form as many vertebrae as your embryo had formed pairs of somites.
Each somite subdivides into four parts: The dermatome --> cells form the inner layer of skin (dermis) The myotome ---> all the skeletal muscle cells of the body The anterior scleroderm --> the posterior half of a vertebra! The posterior scleroderm --> the anterior half of a vertebra!
Not a typographical error! Each somite gives cells to 2 vertebrae
That way muscles run from one vertebra to the next, In his developmental biology textbook, Scott Gilbert advocates the existence of one more sub-part of somites, which he names the "Syndetome", which is postulated to be the cause of formation of tendons. Gilbert admits you can't actually see this structure: "Because there is no obvious morphological distinction between the sclerotome and syndetome cells (they are both mesenchymal), our knowledge...had to wait until we had molecular markers...etc." What this amounts to is inventing a new name for a part of the somites based on discovery that messenger RNA for a certain protein is concentrated there. We all better hope there aren't too many more parts of somites where particular messenger RNAs can be detected, because if there are then somebody is going to invent names for every one of them, like Thisotome and Thatotome! Please notice that this is very different from the theory of how tendons form that is shown in my own photographs and drawings on this web page and with the evidence that David Stopak and I published (some of which is shown on this web page, and more of which will be projected in class, which will include movies of stuff getting pulled around.) The evidence that Stopak and I produced was that mechanical pulling forces occur at certain places in developing embryos, and in tissue culture, and that these forces could line up collagen fibers and also line up muscle cells, and produced muscle-shaped masses of cells, attached at their ends by strands of collagen. We also did experiments injecting collagen from other species into embryos, and finding that it often gets rearranged to be part of tendons. You could say we made artificial tendons and artificial muscles. Contrast this with how some textbooks approach the problem of explaining the mechanism of tendon formation. Some scientists have discovered a protein that occurs in tendons and also occurs in part of somites; they named this protein "scleraxis". They have one beautiful photograph of "in situ" labeling of the locations of the messenger RNA for this protein in a series of stripes apparently in the somites of a chicken embryo. They also have two fluorescence photographs of histological sections through newborn mice on which the DNA base sequence for the Green Fluorescent Protein "had been fused on the scleraxis enhancer". That sounds like it means the GFP base sequence was inserted into a particular part of the mouse genome whose function is to stimulate transcription of the gene for this "scleraxis" protein. Really, what should have been done would have been to attach the GFP base sequence to the actual gene for the scleraxis protein, and not to its enhancer. And I bet that's really what they did. Either way, the interpretation is that scleraxis protein is getting made just at the places where tendons are forming, which is tacitly interpreted to mean that tendons are caused to form by local stimulation of synthesis of this particular protein, at locations that are geometrically between cartilages (which develop from sclerotome, of course) and muscles (which develop from myotome, of course). They even have a diagram of which proteins supposedly stimulate and inhibit each other! The true explanation for the formation of tendons will eventually be discovered, and this explanation will include combinations of gene activation and also mechanical forces, and probably other processes we don't know about yet. Lots of people assume that every body structure forms where it does because some set of genes gets stimulated to be transcribed at that location. No forces are involved; unless forces are too obvious to be ignored, in which case you can say that they are caused by "Thermodynamics" (which is used as a prestigious, pretentious, but meaningless "black box"). Please permit me a small amount of sarcasm: How do elevators work? Elevators work by buttons. You push the button, and that turns on the genes for making an elevator. Then, you get on the elevator, and push another button, which activates genes for the elevator being at another location. Of course, elevator buttons cause stimulation of electric motors up near the roof, etc. But you can't see the motors and the cables; you can just see the buttons. So imagine every time somebody discovers a button, they take that as evidence against the existence of motors and cables; because that's sort of what has happened here. A researcher, Clifford Tabin, whom I know, discovered a protein that gets made in areas where tendons are forming, and also gets made in certain parts of somites. Their methods are in situ hybridization of mRNA and insertion of GFP into genes. So they can "see" where their gene gets transcribed, and also "see" where their protein becomes located. Maybe they also think they can "see" where the enhancer regions are being stimulated? Among the many things that they can't "see" are mechanical forces; so they don't give them any thought. Even though it was proven long ago that skeletal muscle cells differentiate in the somite, and move from there to the arms and legs, nobody worries what moves them to just the right places to be connected by tendons to the skeleton. Stopak and I invented and used methods that could "see" long-range pulling forces; therefore our theories of tendon formation were based on what our methods could "see" (i.e. traction forces). We couldn't see gene transcription; so transcription wasn't explicitly part of our theory; but we did and do believe that some system of gene activation is what controls where mechanical forces become stronger or weaker. Gene control is one part of a causal chain, and mechanical forces are another part of that same causal chain. Textbooks ignore mechanical forces, except for "thermodynamics", which is a "black box" that it doesn't understand at all.
Somites only temporarily exist in the embryo: My own crank theory has been that the physical=mechanical causes of somite separation are key parts of the spacing mechanism, and not just "obeying orders" from Notch genes, etc. Otherwise, why not skip forming somites, and have the genes directly control the locations of vertebrae, ribs, ganglia, etc. |











 |
Three somites Dermatomes farthest to the left Myotomes right under them Then sclerotomes The continuous blue stripe on the right is the neural tube | |
 |
Myotomes in salamander | |
 |
Polarization microscopy shows that myotomes develop muscle fibers very early | |
 |
Alternating anterior and posterior sclerotomes | |
 |
Longitudinal section of 3 somites, that goes right through the myotome |
|
Intermediate mesoderm differentiates to form
kidneys, kidney ducts, and male sex ducts. The most anterior intermediate mesoderm forms the pronephros ("head kidney") (one on each side)
Pronephros
From each pronephros extends a pronephric duct.
Intermediate mesoderm tubules
In amphibia all the rest of the intermediate mesoderm differentiates
into a pair (right & left) of adult kidneys, which use the pronephric duct to carry urine, and to also carry sperm in males.
These adult kidneys are called the metanephros,
and each one has a special duct, called a ureter.
While we are embryos, we use a separate kidney, that differentiates from the middle parts of the intermediate mesoderm. The lateral plate mesoderm splits into two sheets (by its previously-mesenchymal cells changing into epithelial cells, with their apical ends facing each other, creating a fluid space between them.
The space between them is the coelom, or coelomic cavity. The female sex ducts (Oviducts; Fallopian tubules, Uterus) differentiate from outfoldings in the coelomic wall.
|