Lecture notes for Friday, March 11, 2016
Retino-Tectal Nerve Connections
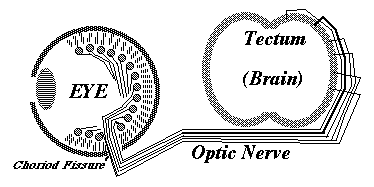
Ganglion cells in the developing eye extend axons toward the "blind spot", where the optic nerve leaves the back of the eye, and where there are no rods and cones, and no vision. Nobody knows (or asks) what guidance mechanism steers the fibers toward the blind spot. But there must be some mechanism.
From the blind spot, the optic nerve growth cones crawl together, side by side, under the brain, passing perpendicularly among nerve fibers coming from the other eye, and up the other side of the brain, eventually spreading out across the upper surface of the midbrain ("the optic tectum"), and forming a neural projection (map-like geometric pattern of connection) on the opposite side of the brain from the side where they originated. Right eye innervates left midbrain; Left eye innervates right midbrain.

-----------------------------------------
a digression on chemotaxis
The affinity could be either selective adhesiveness ("haptotaxis ") or more like chemotaxis.Either of which could be turning of cell locomotion toward increasing chemical concentration, or could be decreasing/increasing of turning, or decreasing vs. increasing of retraction .
Scientists tend to think of chemotaxis as ONE mechanism, But bacterial chemotaxis is by increase in random turning when attractant concentration decreases.
Leucocyte & Dictyostelium chemotaxis is by turning of locomotion toward side where higher concentration of attractant.
Two videos of computer simulations of mechanism of chemotaxis, with audio comments:
-
first video: demonstration of behavioral rules that can produce the effects of chemotaxis
second video: user's choice of different rules
-----------------------------------------
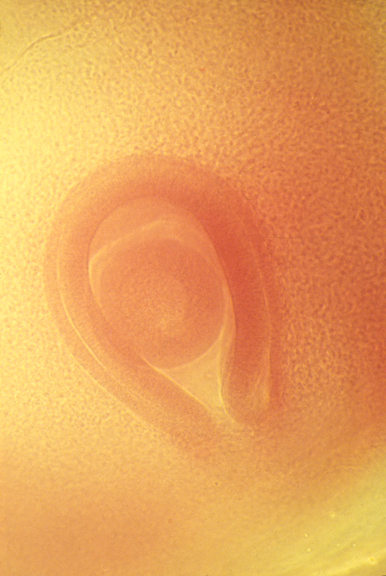
Embryonic eyeball, with retina surrounding it.
The groove is the location of the optic nerve.

Cross-section of developing retina

Cross-section of optic nerve
The "retino-tectal projection " is one of dozens or maybe hundreds of map-like patterns of connection in the human nervous system. We can't be sure they are all caused by the same basic mechanism; but maybe they are, except with differences in adhesion proteins.
Two other examples of neural projections, one of sensory nerves and the other of motor nerves are illustrated by a pair of diagrams in many Psychology textbooks, that shows the whole human body spread out across parts of the cerebrum. Where the diagrams of the face are drawn means those are the parts of the brain that receive nerve impulses from the face (probably relayed via several nerves).
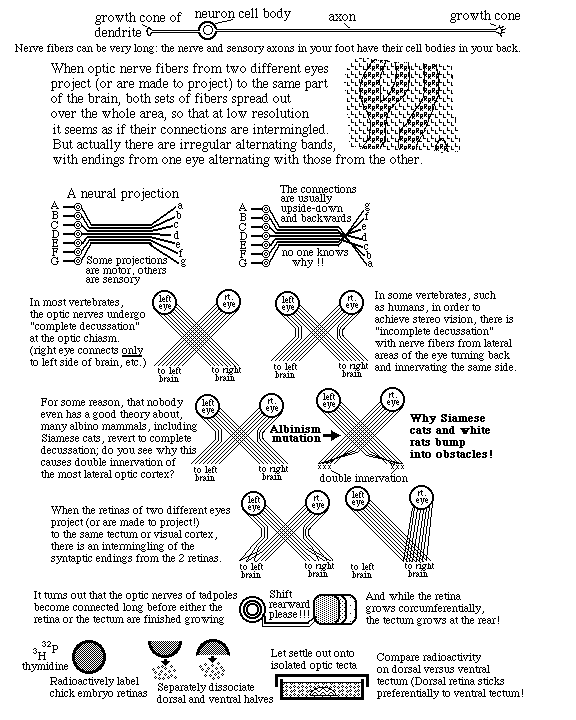
In animals having stereo vision, some nerves from the right eye turn back and innervate the right optic tectum. & some nerves from the left eye turn back and innervate the left optic tectum. This turning back fails to occur in white mice, white rats & Persian cats.
On the tectum roof ("tectum" is Latin for "roof", Spanish word is "Techo") the ganglion cell axon growth cones diverge and form a map-like pattern of the parts of the retina that they came from. Nerves in the front of the retina innervate the rear of the tectum; Nerves at top of retina innervate lateral tectum.
The experiment at the bottom of this figure: Chick embryo eyes were labeled with radioactive isotopes, cut into dorsal and ventral halves and dissociated into individual cells.
Chick embryo midbrains were dissected out and pinned to the bottom of Petri dishes in saline.
Dissociated retinal cells from dorsal retinas were pipetted onto the midbrains.
After a set time, not-yet-attached retinal cells were washed off of the midbrain.
Measurements of radioactivity showed that more dorsal retina cells stuck to lateral midbrain. And more ventral retinal cells stuck to more medial parts of the midbrain. i.e. Retinal cells stick about twice as well to the general part of the brain that axons from there would have innervated.
This was taken as evidence of selective adhesiveness. (And as evidence that neural projections are caused by quantitatively selective adhesiveness) (Prof. Steve Roth, Dept of Biology, U of Penn, was the leader of this research) Vertebrate nervous systems contain hundreds of such map-like connections, which are called "neural projections" A large fraction of these neural projections interconnect upside down and backwards (like the optic nerve).
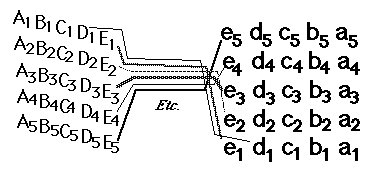
Nerves from the top half of the retina adhere more strongly to the bottom half of the tectum. This was predicted by theories of adhesion gradients, but has turned out to be caused by "ephrin" proteins that stimulate release of cell-cell adhesions.

Nerve axons crawl much farther on molecules from the area of brain they would have innervated.
The more ephrin a cell has on its surface, the less ephrin receptor will be needed to induce retraction.
The less ephrin a nerve cell has on its surface, the higher the concentration of ephrin receptor will be needed to induce detachment and retraction of nerve growth cones.
As an analogy, consider people running across really hot beach sand, toward the water. If people differ in sensitivity to pain, who will stop running closest to the ocean? If there is a gradient of temperature of the sand? Which direction would this sand temperature gradient need to increase?
Some textbooks refer to induced de-adhesion as "repulsion" (which seems misleading to me)
Long range effects of diffusible chemicals have also been discovered ("repulsion" & "attraction")
Plus there are perpendicular gradients of ephrin A and ephrin B, which each have their own specific receptors.