Lecture notes for Wednesday, February 14, 2018
Instabilities that Create Geometric PatternsSomite formation: breaking displacement symmetry
Sequential formation of somites from anterior to posterior in a chick embryo
CLOCK and WAVEFRONT HYPOTHESIS: The "Clock-and-Wavefront Hypothesis" was invented by Cooke and Zeeman to explain somite formation, and especially the ability of embryos to adjust somite size in proportion to the amount of paraxial mesoderm to be subdivided (If you surgically remove tissue, then smaller somites are made). The theory is/was that one quantity oscillates higher and lower in amount, while another variable forms a gradient that gradually increases in amount along its length. A somite is supposed to be split off each time the oscillator increases. Molecules (mostly proteins) have been discovered to behave as the theory predicted. But there are surprises! Three gradients have been found: Fibroblast Growth Factor, Wnt and Retinoic acid (the RA gradient running in the opposite direction [anterior high to posterior low] to the others). Fish use different "clock" substances than birds & mammals (Notch protein) Arthropods also use clocks & wavefronts; which wasn't expected.
REACTION-DIFFUSION SYSTEMS: Turing's & other reaction-diffusion systems "break" displacement symmetry. video with narration showing a pascal program with these rules. This was shown in class without the narration. Please watch the whole thing again with the sound.
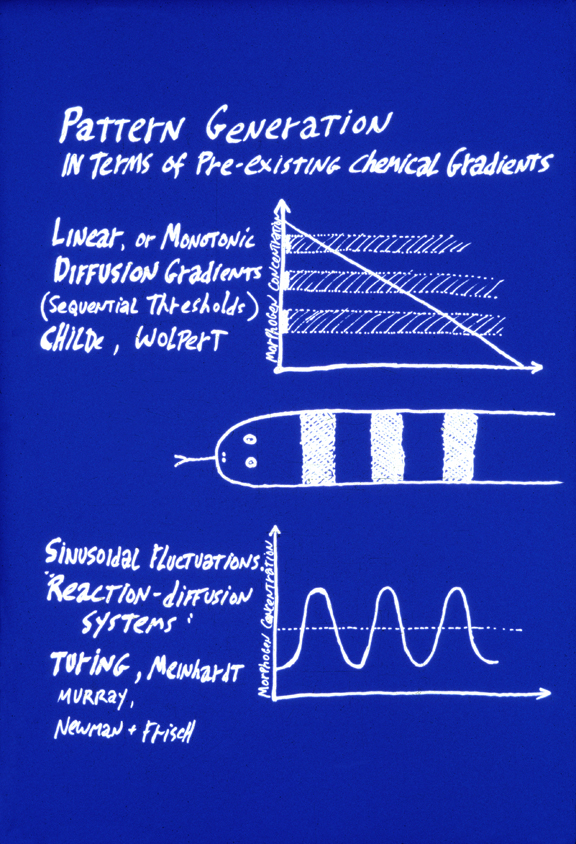
CONTRAST REACTION-DIFFUSION WITH POSITIONAL INFORMATION:
The top of this figure represents the theory of "Positional Information"
This concept was invented by Lewis Wolpert, specifically as a way to explain
b) Each cell measures the three concentrations at its location of these three "morphogen" chemicals. c) An unspecified molecular mechanism "interprets" the combination of three morphogen concentrations.
(For example, if concentration #1 were 22% maximum, and #2 were 55% maximum
Alan Turing's reaction-diffusion system is an alternative way to generate the same striping pattern. In this case, the concentration of some substance (also called a morphogen) varies along
** The chemical morphogens of positional information are very different that what
*** The spatial variation of morphogen concentrations does not form any geometric
**** Most developmental biology textbooks do not distinguish between these uses of
Rules for Turing's mechanism: Please memorize these three simple rules.(Although these are not the only rules that can produce patterns, and they are not even the same as the rules Turing proposed, they are a good example of how simple the rules can be.rule# 1) The concentration of chemical "A" causes more "A" and also more of chemical "B" to be produced. (= Synthesized? Activated? Released from vesicles? Anything!)
rule# 2) Both "A" and "B" are destroyed
(=Inactivated? Reabsorbed? Anything!)
rule# 3) "B"diffuses faster than "A"
The result of obeying these rules is formation of alternating waves of high and low concentration of both A and B.
The peaks of A and B form at the same locations. The wave-length increase in proportion to the ratio of diffusion rates, and also changes if A or B reaction rates are changed. These rules will generate evenly spaced waves, either when initiated by Brownian motion of the chemicals, or by local differences in concentrations of A or B, or other perturbations. In two dimensions these rules will generate peaks or waves (depending on how you initiate them) In three dimensions, they generate regularly-spaced blobs of higher concentrations of A and B. Textbooks tend to assume only irregular patterns can be produced when randomness is used to initiate the process. Computer simulations show this is not true. (A certain textbook has a graph showing the supposed irregularity.)
Another widely-believed fallacy is that B has to diffuse a lot faster than A.
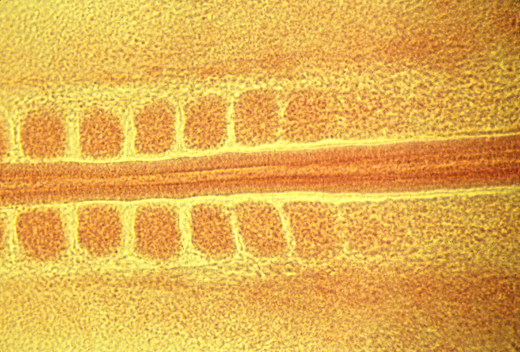
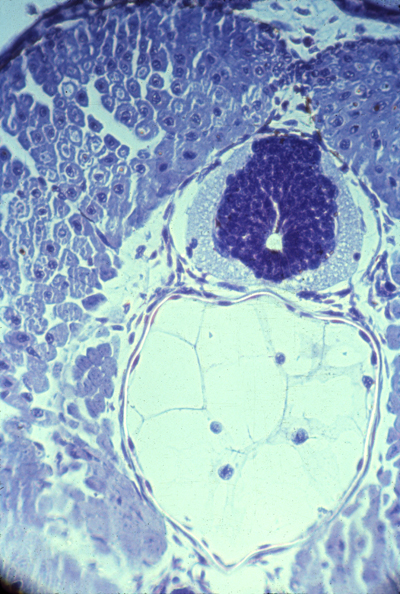
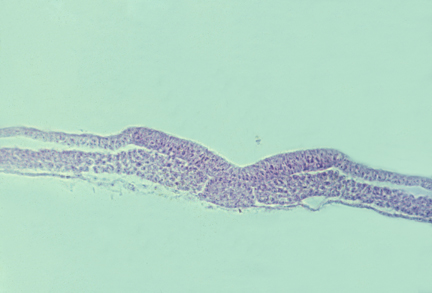
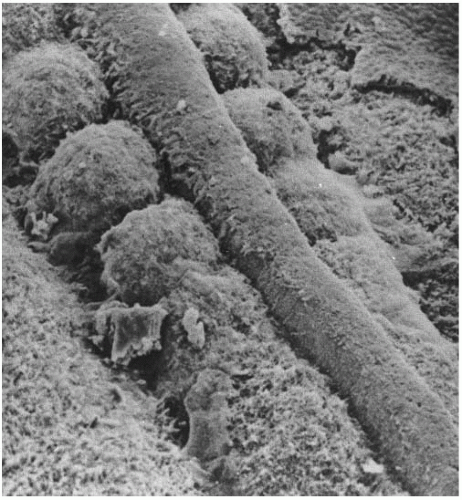
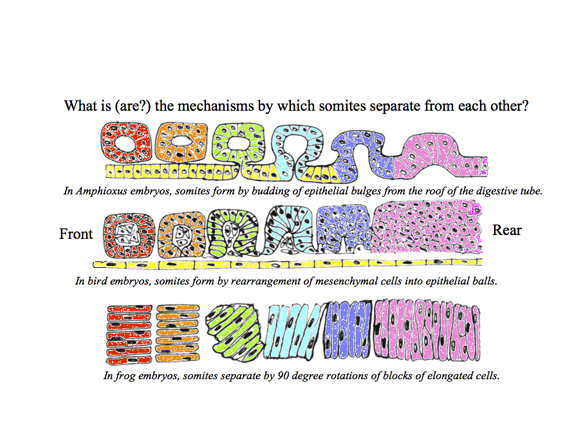
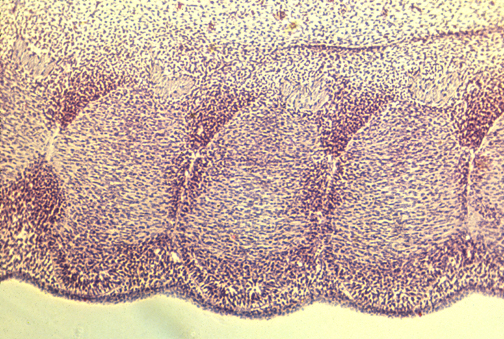
Somites and Notochord
cross-section of an amphibian embryo showing the neural tube (dark)
longitudinal section of a salamander notochord
early neurulation in a chick embryo
scanning electron microscope image of chick embryo somites; by Kathryn Tosney
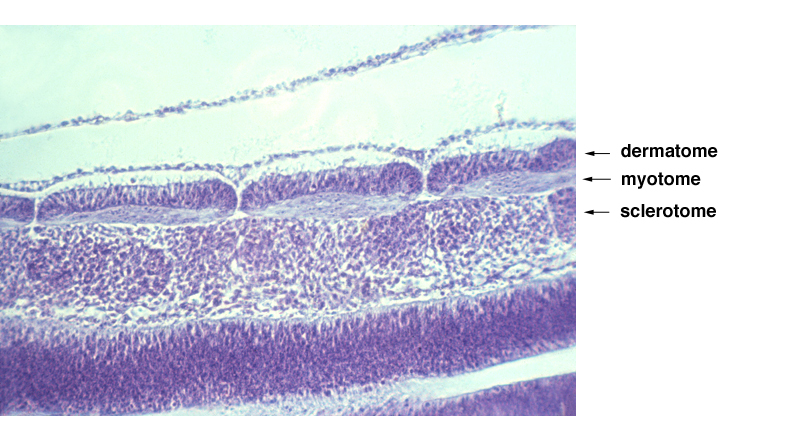
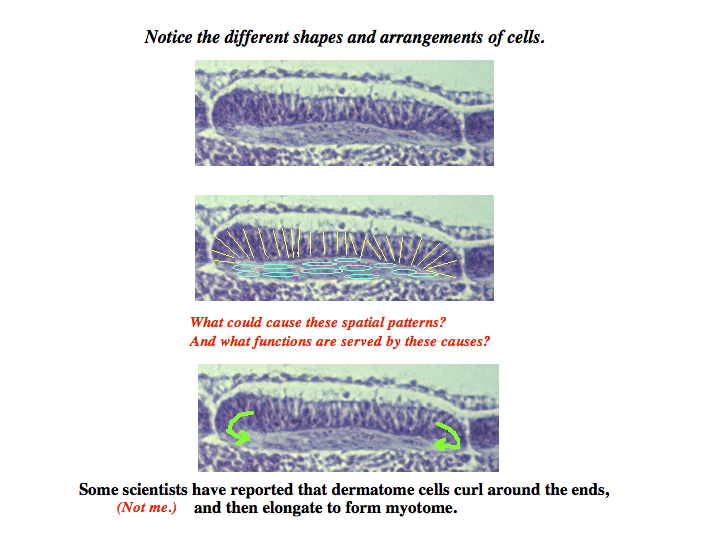
subdivisions of somites: dermatome becomes skin, myotome becomes muscles, sclerotome becomes vertebrae
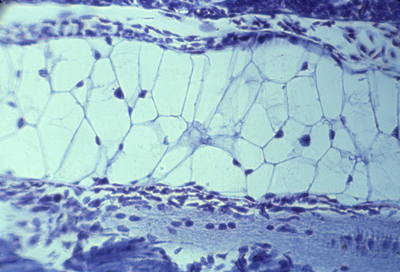
myotome formation
section of myotome
|