Embryology Biology 441 Spring 2011 Albert Harris
Embryology of the Nervous System
Other cells of the central and peripheral cells are called glia, or glial cells.

nerve axons
axons and glia
One example of a kind of glial cell are Schwann cells. These wrap parts of their plasma membrane around nerve axons, to form the myelin sheath (in the peripheral nervous system).
Schwann cells are derived from neural crest cells.
Inside the brain and spinal cord, there are several other kinds of glial cells, such as "oligodendrocytes".
Nerve fibers are long strands of cytoplasm and plasma membrane. The longer nerve fibers are called axons; and most short nerve fibers are called dendrites.
I used to assume that the difference between axons and dendrites must only be size. But it has been discovered that the microtubules inside axons are all oriented with their plus ends in the same direction, away from the cell body; whereas inside dendrites some of the microtubules are oriented with the plus ends away from the cell body, and others oriented in the opposite direction.
Both axons and dendrites are formed by active "amoeboid" crawling locomotion of special areas of cytoplasm at their tips called "growth cones". Axon elongation is a special case of crawling locomotion, not simply growth. Tissue culture was invented by Ross Harrison to prove this fact.
movie showing axon elonagation
"Motor nerves" carry nerve impulses from some part of the central nervous system to muscle cells (or sometimes to gland cells), and stimulate the muscles to contract. The motor nerve cell bodies are located inside your spinal cord.
nerve innervating muscle in tadpole
nerve-motor synapses
Autonomic nerves are a special kind of motor nerves, that control involuntary processes like the rate the heart beats, and intestinal functions
"Sensory nerves" carry nerve impulses from some part of the periphery, back to the spinal cord, or sometimes directly to the brain. Sensory nerve cell bodies are located in a series of spinal sensory ganglia, one pair per somite, next to your spinal cord.
Notice how long the axons have to be that stimulate your foot and hand muscles, and that sense touch, pain etc. in your hands and feet.
These axons have to reach all the way from between your shoulders out to your hands, and all the way from your lower back, down to your feet.
So we are talking about cells that are 2 or 3 feet long, but only a micron, or up to 5 or 10 microns in diameter.
{In case you are interested, those giant axons in squids are multi-nucleate, and formed by fusion of thousands of original cells.}
The neurons in our bodies are all unicellular. Nerve cells never undergo mitosis, or even synthesize DNA, after they have started making an axon.
If some nerve axons are cut in your arm, or leg or back, then the cut end will re-form a "growth cone", which will resume active locomotion, and sometimes find its way all the way back to the original muscle cell, and re-form a synapse or crawl back to the original location in the skin.
But in the embryo, they hadn't needed to crawl nearly as far, to reach their correct connections.
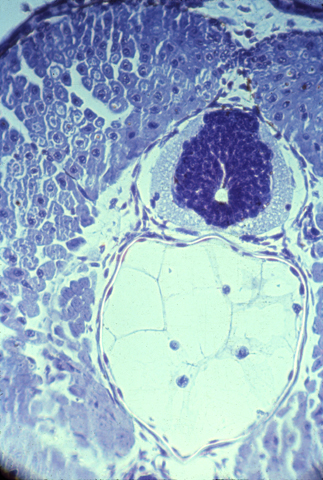
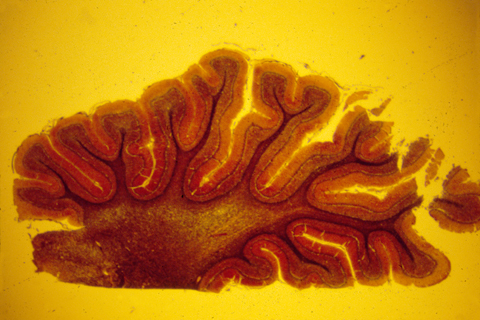
The brain and spinal cord are made of nerve cells, axons, and glial cells tightly packed together. The following photograph shows a histological cross section of the spinal cord of a salamander. The "white matter" is lots of nerve axons packed together. The blue-stained nuclei surrounded by the white matter shows the location of a mass of nerve cell bodies (this is called the "grey matter" because of its grey color when not dyed.
The human central nervous system is also subdivided into grey matter and white matter, except that in brains of higher animals, the grey matter is on the outer surface, and the white matter more interior. The following are histological sections of parts of a mammal brain, stained (dyed) with silver metal in a way that colors only a random 1 or 2 % of the cells, but colors all of the cell body, axons and dendrites of this minority of cells that are stained. Without this method, the cellular structure of the central nervous system would never have been mapped. That is because the nerve cells and fibers are packed together so tightly that staining them all would produce a solid mass of color.

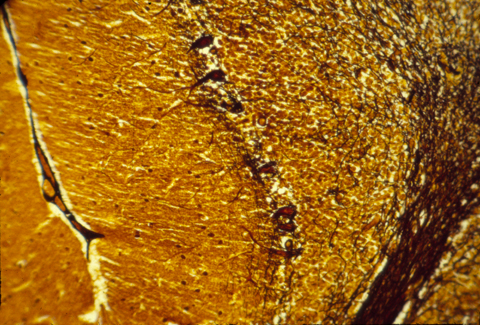
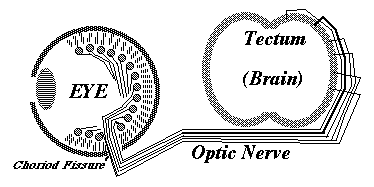
Much research has been concentrated in the mechanisms that guide the growth cones of the developing optic nerve from the neural retina of the eye, back to the particular part of the brain that it innervates. The retinal cells whose axons form the optic nerve are called "ganglion cells", you have about a million of them in each eye, and each one sends of single axon back to the opposite side of the brain (mostly!).
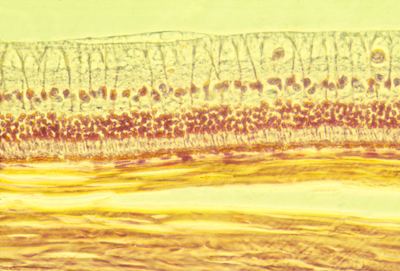
Histological section of the retina of a rabbit. The light from the pupil and lens comes from the top (as this section is oriented), passes through the optic nerve axons at the top, and the large ganglion cell nuclei below and among them, and on downward to the rod and cone cells in the middle. Below then is the pigmented retina, and below that you can see thick strong sheets of collagen fibers that are wrapped around the eyeball and give it strength.
In birds, amphibians and fish, the part of the brain they innervate is called the "optic tectum". At the tectum, the growth cones of the optic nerve axons spread out over the brain surface is the same geometric pattern as the parts of the retina where they are coming from.
They then form synapses in what amounts to an up-side down and backwards (reversed back to front) "map" of the parts of the retina where their ganglion cells are located. This pattern of connection is called a neural projection.
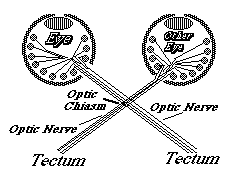
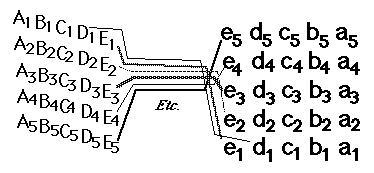 The retino-tectal projection is one among dozens, or probably hundreds of different neural projections in our nervous system. For example, the sensory nerves of the whole surface of the body form a neural projection to a strip along the roof of the forebrain;
And the motor nerves for all the voluntary muscles of the body form a neural projection from another strip of the roof of the forebrain, side by side with the sensory projection,
separated by a certain infolding of the roof of the brain.
The retino-tectal projection is one among dozens, or probably hundreds of different neural projections in our nervous system. For example, the sensory nerves of the whole surface of the body form a neural projection to a strip along the roof of the forebrain;
And the motor nerves for all the voluntary muscles of the body form a neural projection from another strip of the roof of the forebrain, side by side with the sensory projection,
separated by a certain infolding of the roof of the brain.
The causal mechanism for the formation of the retinotectal connection has long been thought to be some kind of adhesive specificity. The first demonstration of this was about 35 years ago.
The current belief is that special proteins called ephrins occur in tectal cells, where they vary in number per cell according to a gradient (notice this is not a diffusion gradient). Another kind of cell surface receptor, called an ephrin receptor, is present on the surfaces of the ganglion cells and their axons. Ephrin proteins bind selectively to ephrin receptor proteins; and the higher the concentration of ephrins, then the lower the concentration of ephrin receptors needed to produce an the essential effect - but this effect (surprise!) is not adhesion between axon and brain cell, but somewhat the reverse!
Ephrin binding inhibits further locomotion of a growth cone, often resulting in its retraction a short distance. The basic idea is that this is super-imposed on some less specific attraction of optic nerve growth cones, with inhibition of crawling being produced by ephrin-ephrin receptor binding. When the farthest part of the tectum has he higher concentration of ephrin, then only the axons with the lowest concentrations of ephrin receptors will be able to get that far before further crawling is blocked. Axons with the highest concentrations of ephrin receptors will be stopped at the lower side of the tectum, where the brain cells have the highest concentrations of ephrin.
There are two classes of ephrins and receptors, ephrin A and ephrin B, and the evidence is that one controls the dorso-ventral axis and the other controls the anterior-posterior axis. There is still much to be learned however.
Some other kinds of evidence suggests that ganglion cell axons interact with each other. For example, Martha Constantine Paton did experiments in which an extra eye was grafted onto the side a frog head, and the pattern of innervation by their ganglion cells was studied in the tectum. Instead of the nerves from both eyes connecting in a homogeneous mixture, they formed stripes. Each stripe of the tectal surface became innervated mostly by axons from one or other of the eyes, and the adjacent stripes on either side of that stripe became innervated mostly by axons from the other eye.

GREETINGS, EARTH PEOPLE
There are several different forms of chemotaxis.
The word chemotaxis used to refer to several, logically distinct phenomena.In no case of chemotaxis does a chemical gradient exert a physical pull, or push.
(The smell of good food does not pull on your nose, in any physical sense; nor does a bad smell push you away from it.)
Nevertheless, textbooks and others often talk about chemotaxis "attracting" cells, so sometimes readers get the wrong idea.
Phenomena that various people call chemotaxis:
#1) If cells keep going in straight lines as long as they detect increases in the chemotactic "attractant" substance (now versus previously), but turn in some (random) direction if they detect decreases in concentration.#1 ½ ) If cells move approximately straight, compare the concentration at the front relative to the back, & don't turn unless the rear gets to be more.
1 ¾ ) Concentrations at growth cones could be compared with concentrations farther back along the axon, and extension could continue if the concentration is higher at the growth cone, but turn otherwise.
1 & 7/8 ) Compare concentrations at tips and bases of filopodia...
#2) If cells compare the local concentration of attractant substance on their right side versus their left side, and turn right when the concentration of attractant is higher on their right side, and turn left when that concentration is higher on their left side.
#2 ½) Or compare concentrations detected by filopodia on right versus left, etc.
#3) If the concentration of attractant is measured at many locations all around each cell's margins, and a front end is induced to form at whichever part of the cell margin detects the highest concentration.
3 ½) Use filopodia to compare concentrations.
#4) Slow down or stop where the attractant concentration becomes higher than some threshold amount. (this can look surprisingly much like attraction) Really it's a form of "trap action".
#4 ½) Slow down where the attractant concentration is equally large on all sides of the cell...
#4 ¾) Or slow down where the attractant concentration has greatly decreased its former rate of increase. Or turn?
#5) ? Can you invent more? They exist! Concepts like this have technical uses.
Bacteria and ciliate protozoa (& sperm?) use #1 from this list.
Polymorphonuclear leucocytes use #2.
Dictyostelium use #3.
Nerves probably use #3 ½, but it's hard to be sure.
Do you think all these phenomena deserve to be called by the same name?
Some purists think we should call #1 "chemokinesis" And reserve the word "chemotaxis" for #2 and/or #3.
They have a good point. But the word chemotaxis was first invented by scientists studying bacterial chemotaxis, so that creates priority.
There are also many examples of "negative chemotaxis", Which means that cells move toward lower concentrations of something.
Vocabulary should help us understand how things work, not cause us to equate phenomena with fundamentally different causes, and also not let words substitute for information.
"These cells move by chemotaxis...
"Those airplanes move by radar...
"Those ships move by sonar... or are pushed by periscopes.
_____________________________________________________________
Haptotaxis means cell movement directed in response to a gradient of adhesiveness.
The discoverer of haptotaxis literally believed that crawling cells get pulled
along by increased adhesiveness of gradients of metal thickness etc.
That conclusion was published in Nature, and seemed hard to disprove, at the time.
Evidence eventually persuaded him that movement of cells up adhesion gradients results from a tug of war between different sides of individual cells, each pulling in a different direction, so that the side of each cell where the cell-substratum adhesion is strongest "wins", and becomes the front.
* If a cell is put on a small adhesive island, its opposite sides will crawl right off the island, thus decreasing total adhesiveness.
* Those kinds of leucocytes which have one distinct "front" do not orient their locomotion to adhesion gradients. (There is no tug of war, for them.)
* You might be able to think of other experiments, or observations.
In order for the "Differential Adhesion Hypothesis" to work, cells would literally need to be pulled by the force of adhesion, as if adhesive tape were to pull things like a magnet does.
Force is often needed to break adhesions, but the formation of adhesions does not itself exert a pulling force.
"Thermodynamics" can be used as a black box to intimidate people, so that they will accept silly conclusions for fear of seeming ignorant.
One important reason to take courses in thermodynamics is to immunize yourself against misuse of this very useful subject.
________________________________________________________________
Neural Projections are caused by what amounts to negative haptotaxis.
Imagine 100 people running toward the ocean over hot sand.
& with a temperature gradient - hottest sand nearer the ocean,
and the people vary in tolerance for pain,
or maybe toughness of their feet.
Everyone will run as far as they can stand the pain, & then sit down.
Those who can stand less heat stop farthest from the ocean. So quantitative differences are converted into geometric locations.
____________________________________________________________
Imagine a concentration gradient of a chemical that induces cells to retract, plus cells with differing amounts of receptors to this chemical.
The chemical is like the hot sand on the beach... The receptor is like the people's pain sensitivity.
Proteins called "Ephrins" bind to "Ephrin Receptors" on cell surfaces.
There are several kinds of ephrins: Ephrin A, Ephrin B
And cells with ephrin A receptors retract when they encounter too much ephrin A
Binding of "Ephrin" proteins to their specific "Ephrin Receptor" proteins induces cells to detach their adhesions, and retract backward away from them.
Ephrin receptors are tyrosine kinases, that induce some change either in cell adhesiveness, or possibly contractility.
This is a very important way of guiding cells, and especially nerve growth cones.
The Gilbert textbook refers to this process as pushing (speaking figuratively), But please do not think that ephrin really pushes
Gradients of concentrations of ephrin A (somehow fixed in place!) exist on the surface of the optic tectum.
Optic nerve growth cones have ephrin receptors on their surfaces; And there is a sideways gradient in the amounts of these receptors (i.e. more on the nerves from one side of the retina, less on the nerves from the middle, and least on the cells from the other side of the retina.)
The nerves with the smallest amount of receptors can continue to crawl farthest across the tectum, from what we can call the "less ephrin side" toward the more ephrin side.
(Nobody actually calls them that; I am trying to make it easier to visualize.)
These discoveries continue, and are the culmination of a long line of research going back to tests whether dissociated retinal cells would adhere more strongly to the side of the tectum that their axons would innervate.
Please notice:
1) One gradient is needed in the anterior-posterior axis and a different pair of receptors and binding molecules are needed in the dorso-ventral axis. [The latter is supposed to use one of the wnt proteins]
2) None of these gradients are diffusion gradients. What causes the gradients to form is another, also interesting, question.
3) Some additional directional mechanism is needed to cause the nerve growth cones to keep trying to go up the gradient that breaks adhesions. (analogous to barefoot people running toward the ocean).
4) None of the gradients push or pull, in any literal sense. (But rather favoring or thwarting active locomotion of nerve growth cones.]
5) Computer simulations might help visualize this, and help design experiments.
6) And please consider the following puzzling experiments, in which halves or retinas or halves of tecta are removed before innervation.
________________________________________________________________
A somewhat surprising result is what happens when half a retina is destroyed...
Would you expect the remaining optic nerve fibers to innervate only half the tectum? Or might these nerve fibers spread out their connections over the whole tectum?
Or when half a tectum is destroyed...
What are the two main possibilities? Where would the optic nerve growth cones grow?
The same place as usual? With half having nowhere to go? Or "move down, make room for everybody"?
Or when the optic nerves from two eyes are guided so that they can't help innervation the same tectum?
What alternative possible results can you think of?
Maybe all the nerves from one eye form a projection over on one part of the tectum, and those from the other eye form another projection next to it?
Or all the optic nerve growth cones mix randomly?
Or maybe they alternate in stripes...? (But that would be silly! Or would it?)
__________________________________________________________________
Neural connections were traced (never mind how) from right or left eyes back to where they innervate the brain (not the tectum, in the first experiments) and their connections were mixed with those from the other eye, in geometric patterns approximately like the one below. ("R" means a connection from the right eye; guess what "L" means!)
RRRLLRRRRLLLRRLLLRRRLLLRRRLLLRRLLRRLLRRRLLLRRLLRR RRRLLLRRRLLLRRLLLRRRLLLRRRRLLRRLLRRLLRRLLLRRLLRRR RRRLLLRRRLLLRRLLLRRRRLLLRRLLRRRLLRRLLRRLLRRRLLRRR RRRRRLLRRLLRRRLLLRRRLLLRRRLLRRRLLRRLLRRLLRRRLLLRR RRRRRLLRRLLRRRLLLRRRLLLRRRLLRRLLLRRLLRRLLRRLLLRRR RRRRRLLLRRLLRRRLLLRRLLLRRLLLRRLLLRLLLRRLLRRLLLRRR RRRRRLLLRRLLLRRLLRRRRLLRRLLLLRRLLLRLLLRLLRRLLLRRR RRRRRLLLRRRLLLRLLRRRLLLRRLLRLRLLLRLRLLRLLRRLLLRRR RRRRRLLLRRRLLLRRLLRRLLLRRLLRLLLRLLLRLLLRLLRRLLLRR RRRRRLLLRRRRLLRRLLRRLLLRRLRRLLRRLLRRLLRRLLRRLLRRR RRRRRLLLLRRRLLRRLLLRRLLLRLRRLLRRLLRRLLRRLLRRLLRRR RRRRLLRRLLRRLLLRRLLRRLLLRLRRLLRRLLRRLLRRLLRRRLLLR RRRRLLRRLLRRLLLRRLLRRLLLRLLRRLLRLLRRLLRRLLLRRLLRR RRRRLLRRLLRRLLLRRLLRLLLLRLLRRLLRRLLRLLRRRLLLRLLLR RRRRLLRRLLLRRLLRRLLRLLRLLLRRLLLRRLLRRLLRRLLLRRLLR RRRLLLRRRLLRRLLRRRLLLRRLLRRRLLLRRLLLRLLRRLLRRRLLR RRRLLLRRRLLLRRLLRRLLLRRLLRRRLLRRRLLRRLLRRLLLRRLLR RRRLLRRRRLLRRRLLLRRLLLRRLLRRLLRRLLLRRLLRRLLRRLLLR RRRLLRRRLLLRRRLLLRRRLLRRLLRRLLRRLLRRLLLRRLLLRLLLR RRRLLRRRLLLRRRLLLRRLLLRRLLRRRLLRLLRRLLLRRLLLRLLLR RRRLLLRRLLLRRRLLRRRLLLRRLLRRLLRRLLRRLLLRRLLRRLLLR RRRLLRRRLLLRRRLLLRRLLLRRLLRRLLRRLLRLLLRRRLLRRLLLR RRRLLRRRLLRRRRLLLRRLLRRLLLRRLLRRLLRLLLLRRLLRRLLLR RRRLLRRRLLRRRLLLRRRLLRRLLRRRLLRRLLRLLRLLRLLLRLLRR RRRLLLRRLLRRRLLLRRRLLLRRLLRRLLRLLLLLRRRLLRLLLLRRR RRRLLLRRLLRRRLLLRRRLLLRLLRRLLLRLRLLRRRLLRRLLLRRRR RRRLLLRRLLLRRRLLLRRLLRRLLRRLLRRLRRLLRRLLRRRLLLRRRNotice that the distribution isn't even, and it also isn't random.
It's more like zebra stripes or finger-prints.
In later experiments, both of a frog's optic nerves were forced to innervate one of the frog's optic tecta, and they also formed stripes like this.
Notice that tecta is the plural of tectum.
( Second declension neuter. Latin students rule..)
When trying to figure out what sort of mechanism would produce stripes think about two classes worth of students, each student reeling out string and told to sit as much as possible in the same relative positions relative to their previous neighbors, and competing for seats with students from an entirely different class, who are also trying to sit in the same relative part of the room as previously, and next to the same other students.
Hint: Suppose you have to choose between sitting next to your previous neighbors on your sides, or behind and in front of the students you previously were behind and in front of.
Either preference will tend to produce stripes, it seems to me.