Pattern Formation
CLOCK and WAVEFRONT HYPOTHESIS:The "Clock-and-Wavefront Hypothesis" was invented by Cooke and Zeeman to explain somite formation, and especially the ability of embryos to adjust somite size in proportion to the amount of paraxial mesoderm to be subdivided (If you surgically remove tissue, then smaller somites are made). The theory is/was that one quantity oscillates higher and lower in amount, while another variable forms a gradient that gradually increases in amount along its length. A somite is supposed to be split off each time the oscillator increases.
Molecules (mostly proteins) have been discovered to behave as the theory predicted. But there are surprises!
Three gradients have been found: Fibroblast Growth Factor, Wnt and Retinoic acid (the RA gradient running in the opposite direction [anterior high to posterior low] to the others).
Fish use different "clock" substances than birds & mammals (Notch protein).
Arthropods also use clocks & wavefronts; which wasn't expected.
POSITIONAL INFORMATION:
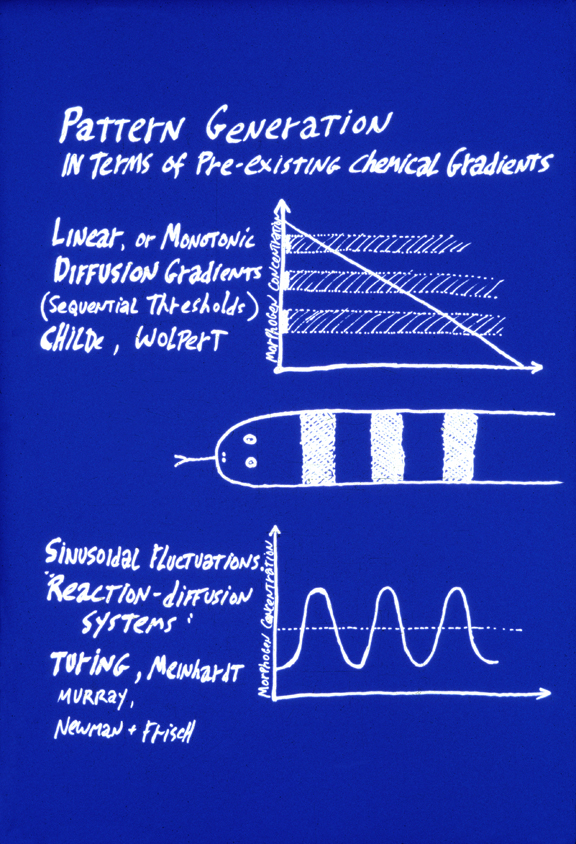
The top of this figure represents the theory of "Positional Information"
This concept was invented by Lewis Wolpert, specifically as a way to explain
the Driesch phenomenon.
e.g. separated 2 cell stage echinoderms developing into
half-size larvae, and all that.
-
a) Three or more linear diffusion gradients form, each perpendicular to the other two.
b) Each cell measures the three concentrations at its location of these three "morphogen" chemicals.
c) An unspecified molecular mechanism "interprets" the combination of three morphogen concentrations.
(For example, if concentration #1 were 22% maximum, and #2 were 55% maximum and #2 were 77% maximum, then the unspecified interpretation could be "differentiate into a chondrocyte".
The key idea is that diffusion gradients should be twice as steep in half-sized embryos, ten times as steep in tenth-sized embryos, half as steep in double-sized embryos.
In my opinion, Wolpert has drawn a conclusion based on plausibility but not much else.
This theory can explain Driesch's observations of regulation, but can't explain Roux's hot needle experiment. Wolpert's "French Flag Model" was also mentioned in class, but no slides were shown. For more information, please see Wikipedia article>
REACTION-DIFFUSION:
Alan Turing's reaction-diffusion system is an alternative way to generate the same striping pattern. In this case, the concentration of some substance (also called a morphogen) varies along the length of the animal in a regular manner, creating a "pre-pattern" for the
eventual stripes in the animal. "
(This wasn't discussed at length in class.)
***************************************
A slide was shown of two figures from the following paper: Kendall, D.G. (1971). Construction of maps from "odd bits of information." Nature 231: 158-159. doi: 10.1038/231158a0. A computer program was used to construct a map of France from information on whether or not each governmental department shared a common boundary with each of the others. The computer-generated map is compared with the actual map of the boundary positions.
LIMB GRAFTS
Triple-branching of 180-degree-rotated limb grafts is an example of diffusion-like filling-in of intermediate behaviors of cells.
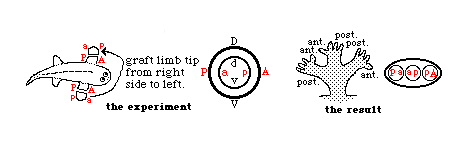
Experiment by Susan Bryant and her collaborators
Another drawing making the same point:
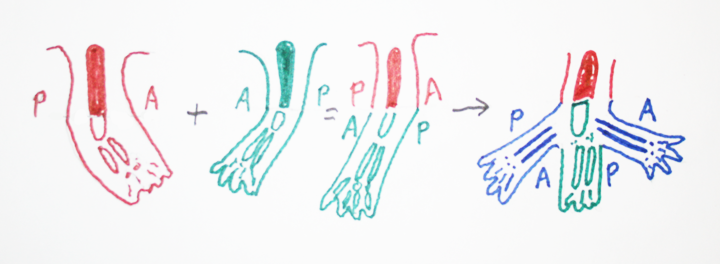
What has happened?
original limb ABCDEFGHIJKLrotated graft: ABCD/HGFE/IJKL
"regeneration" ABCDefgHGFEfghIJKL
But H is happy being between two Gs, one of them newly made.
(Even if H apparently doesn't care not having any nearby "i".)
This must be telling us something profound about how regeneration works.
Being next to a "wrong" neighbor is what gets fixed.
(by replicating extra mirror-image duplicates of "correct" neighbor.)
But tissues are NOT disturbed by absence of a normal neighbor.
They hate discontinuity! But they don't mind duplication!!
This must be telling us something about underlying control mechanisms.
(But it sure isn't easy to put into words whatever it's telling us.)
One question is how cells detect being next to "wrong" neighbors..
Maybe "wrongness" consists of which hox genes you and they transcribe?
Suppose each cell "knows" which hox genes it is itself transcribing,
and also detects which hox genes are being transcribed by nearby cells.
You are happy when your neighbors transcribe hox genes that are adjacent on your chromosomes to the hox genes that you, yourself are transcribing.
But it's intolerable for nearby cells not to transcribe hox genes whose positions on your chromosomes are close to whatever hox genes you are transcribing. (Discontinuity!)
I hope you will think about these issues, but not worry if you can't devise answers.
One final note: Genetic linkage was discovered years before Morgan figured out that its explanation was the relative positions of genes on the chromosomes.