Lecture notes for October 21, 2020
"Embryonic Regulation"
Hans Driesch: discovery of "Regulative development" (in the 1890s). He separated the first 2, and the first 4 cells of starfish embryos, and observed their development into two half-size plutei, or 4 quarter-size. Conversely, you can push two one-cell stages together and sometimes they will develop into double-sized plutei.
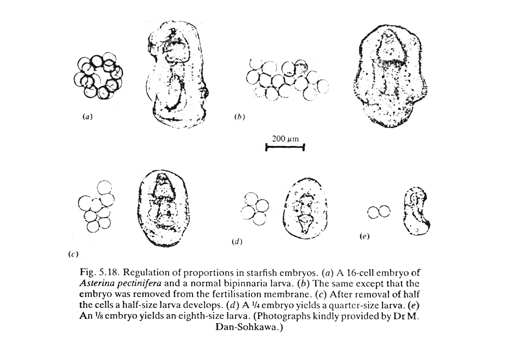
This disproved the theory of different parts of embryos receiving different genes.
But even today scientists don't really understand the mechanism that changes the sizes of parts of embryos in proportion to the size of the whole embryo.

Please don't be misled by the word "regulation",
which can seem to imply some exterior control telling somebody what to do.
This kind of regulation is internal adjustment, to fix damage or overcome abnormality or effects of some external disturbance.
Whatever adjustments cause regulation of shape and adjustment of proportionality of sizes, sometimes require time to adjust.
"Regulative" development contrasts with "mosaic" development, in which cell fates are pre-determined, for example in the nematode Caenorhabditis elegans.
Driesch concluded that embryos are somewhat supernatural
in the sense of being controlled by something mind-like.
----------------
An even better example of size adjustment of parts in proportion to wholes is Dictyostelium formation of fruiting bodies, stalks, and length/width ratios of slime mold slugs.
[See the web notes on amoeboid locomotion and on chemotaxis for more about the life cycle of Dictyostelium. Remember that when they run out of bacteria, they aggregate by chemotaxis, and form multicellular "slugs" that can crawl around. The slugs vary widely in size, depending on how many individual amoebae aggregated to form them. They then differentiate into stationary "fruiting bodies" that consist of a tapering stalk, with a lemon-shaped mass of spores at the top, and have the same proportions over a size range of at least 500.]
If you cut a slug into three segments, all three will develop stalks and fruiting bodies, but the proportions of stalk and fruiting bodies will be somewhat different from each other. Even tiny pieces of a single slug will do this. The image below is a still frame from John Tyler Bonner's movie of the Dictyostelium life cycles and experiments.
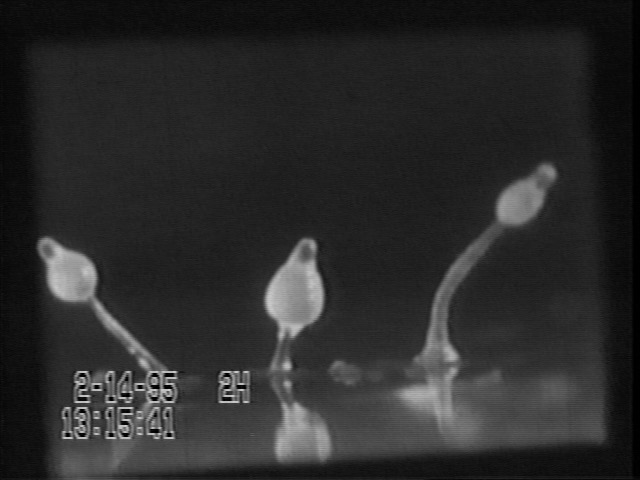
This is even more than Driesch's revolutionary discovery that starfish embryos can "scale" over a sixteen-fold size range.
Note that the slugs bend away from each other. This may be an example of negative chemotaxis.
You can cut a Dictyostelium "slug" into 10 or 100 pieces, and the cells of each fragment will reorganize to form a normally-proportioned slug, and then a scale model stalk and spore mass, one one-hundredth the size the original "slug" would have formed. Echinoderm plutei can develop "scale models" over a volume ratio of 8 to 1 or sometimes 16 to one.
Scaling may have different mechanism in plutei than in slime mold fruiting bodies.
But I regard this as one of the 3 or 4 deepest questions in biology.
What does the ability "to scale" tell us about normal mechanisms that cause
organ formation?
That's the big question. There might be several answers, or one.
Also note that mammal embryos can be separated fairly late (20+ cells) and still develop into normal animals.
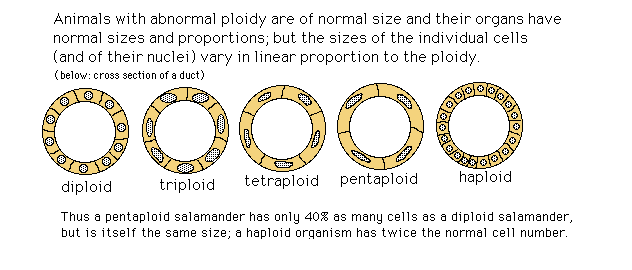
Ploidy
Two interesting aspects of regulative development:1) Tetraploid cells have (exactly!) twice the volume of diploid cells; and haploid cells have exactly half the volume.
2) Organ sizes remain the same, despite cell sizes being half or twice as big as in diploid animals of that species.