Embryology Biology 441 Spring 2011 Albert Harris
Tissues and organs that develop from mesoderm
| Lateral plate mesoderm |
Intermediate mesoderm |
Somites | Notochord | Somites | Intermediate mesoderm |
Lateral plate mesoderm |


|
We are members of the phylum "chordata", all of the members of which have a notochord at some stage of their development. In mammals and birds, the function of the notochord is replaced by the vertebral column (bones of the back-bone+intervertebral discs) But tadpoles and larval fish actually depend on the notochord to allow them to swim by alternating contraction of muscles along each side.
The notochord mesoderm also induces formation of the neural tube. The structure of the notochord is a long stack of (not always) flattened cells, that often have very much the geometry of a stack of coins. Wrapped tightly around the surface of this long cylinder of cells are layers of fibers of type I collagen in many spiraling layers, that alternate in direction (a clockwise layer, then a counterclockwise layer, then another clockwise layer, etc. etc. etc.) The cells inside the notochord then form large vacuoles in their cytoplasm, which makes each cell swell in volume, pushing against the spiral layers of (strong & non-stretchable!) collagen fibers. The result is that the notochord can bend side to side, but can't be compressed along its anterior-posterior axis.
Tail of a living tadpole. The broad line just below the center
Neural tube and notochord
Notochord of salamander
Section through chick embryo
Several odd facts about the notochord: In reptiles, birds and mammals, the notochord cells are the last to leave the surface (epiblast) at the Hensen's Node.
* In some species (humans & turtles), Hensen's node is partly an invagination, & a hollow tube runs down the middle of the notochord * In sections of amphibian embryos, you can see pigment granules tend to accumulate down the middle of the notochord. * The spiral layers of collagen fibers that are the sheath of the notochord are oriented at 90 degrees to each other. This is one of several examples in which collagen fibers mysteriously become oriented in alternating layers, with the fibers of each layer exactly perpendicular to the fibers on the layers above and below.
Also, there are many examples in which muscle cells become lined up in perpendicular alternating layers: Mammal tongues, elephant trunks,
squid tentacles, and in Hydra, both the body and the tentacles.
Polarization micrograph of a section through a mouse tongue
Anybody who could figure out the mechanism that creates these perpendicular layer patterns, and prove their theory true, would go down in biological history. Somites are segmental blocks that form beside the neural tube. At first, there are continuous columns of "paraxial" mesoderm
The drawing above shows the geometrical rearrangements that some scientists have reported to casue somite segmentation in different classes of chordates. Top: Amphioxus, Middle: bird and mammal, Bottom: frogs. Anterior is to right side.
Then these columns spontaneously split apart, one pair at a time.
Longitudinal section of notochord and somites in a developing frog embryo
Somites forming in a fixed whole mount chicken embryo. Anterior to left.
Somites forming in a living chicken embryo. Anterior to left.
The physical mechanism of splitting apart could be active constriction, or could be decreases in cell-cell adhesion proteins. In scanning EM photos, "somitomeres" can be seen, where a somite is going to form, apparently as the first stages of somite formation.
In most species, somites start as hollow epithelial balls,
and conversion of cells from being mesenchymal to epithelial
therefore seems to be part or the process of separation.
Much research has been done on the genes needed for somite formation The most popular categories of theory are 1) That somite segmentation has the same mechanism as formation of segments in flies, using genes analogous to "even-skipped" etc. 2) The "Clock-and-Wavefront Hypothesis", according to which one quantity oscillates higher and lower in amount, while another variable forms a gradient that gradually increases in amount along its length. A somite is supposed to be split off each time the oscillator increases. Really, no one even knows whether we should think of the splits as being the real entities, as opposed to the somites themselves. Links to papers on somite formation, including these theories
For most species, the same number of somites forms in each embryo. You form as many vertebrae as your embryo had formed pairs of somites.
Each somite subdivides into four parts: The dermatome --> cells form the inner layer of skin (dermis) The myotome ---> all the skeletal muscle cells of the body The anterior scleroderm --> the posterior half of a vertebra! The posterior scleroderm --> the anterior half of a vertebra!
Not a typographical error! Each somite gives cells to 2 vertebrae
That way muscles run from one vertebra to the next, On page 459 of the eighth edition of the Gilbert textbook, in figure 14.19, he advocates the existence of one more sub-part of somites, which he names the "Syndetome". Certain researchers have postulated that such a thing exists, and that it is the cause of formation of tendons. Gilbert admits you can't actually see this structure: "Because there is no obvious morphological distinction between the sclerotome and syndetome cells (they are both mesenchymal), our knowledge...had to wait until we had molecular markers...etc." What this amounts to is inventing a new name for a part of the somites based on discovery that messenger RNA for a certain protein is concentrated there. We all better hope there aren't too many more parts of somites where particular messenger RNAs can be detected, because if there are then somebody is going to invent names for every one of them, like Thisotome and Thatotome!
Somites only temporarily exist in the embryo:
|






 |
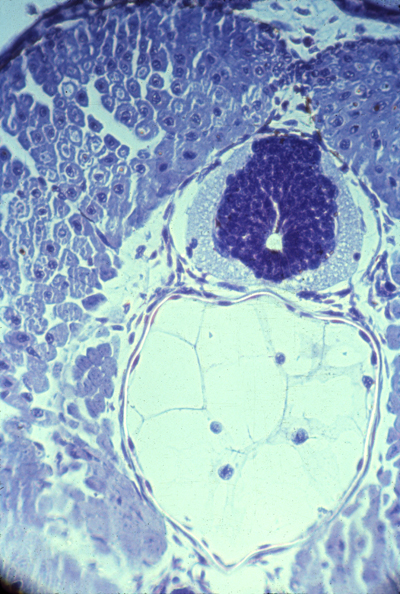
Three somites Dermatomes farthest to the left Myotomes right under them Then sclerotomes The continuous blue stripe on the right is the neural tube | |
 |
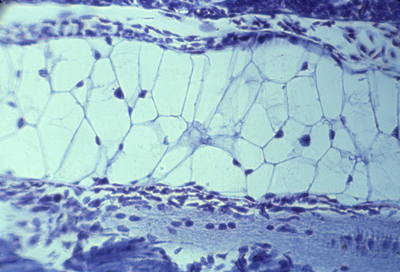
Myotomes in salamander | |
 |
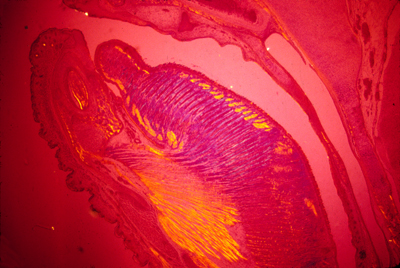
Polarization microscopy shows that myotomes develop muscle fibers very early | |
 |
Alternating anterior and posterior sclerotomes | |
 |
Longitudinal section of 3 somites, that goes right through the myotome |
|
Intermediate mesoderm differentiates to form
kidneys, kidney ducts, and male sex ducts. The most anterior intermediate mesoderm forms the pronephros ("head kidney") (one on each side)
Pronephros
From each pronephros extends a pronephric duct.
Intermediate mesoderm tubules
In amphibia all the rest of the intermediate mesoderm differentiates
into a pair (right & left) of adult kidneys, which use the pronephric duct to carry urine, and to also carry sperm in males.
These adult kidneys are called the metanephros,
and each one has a special duct, called a ureter.
While we are embryos, we use a separate kidney, that differentiates from the middle parts of the intermediate mesoderm. The lateral plate mesoderm splits into two sheets (by its previously-mesenchymal cells changing into epithelial cells, with their apical ends facing each other, creating a fluid space between them.
The space between them is the coelom, or coelomic cavity. The female sex ducts (Oviducts; Fallopian tubules, Uterus) differentiate from outfoldings in the coelomic wall.
|