March 3 lecture
Control Mechanisms & Ectoderm
Somatic Ectoderm:
Hairs each consist of regularly arranged epithelial cells.

Each hair in the skin of every mammal has a mesodermal papilla at its root.

None of the papilla cells forms any part of a hair; They stimulate differentiation of ectodermal cells into hair cells, instead of allowing them to be ordinary skin cells.
If the papillae are large, the diameter of the hair will be thick;. If the papillae are small, then the diameter of the hair will be narrow; If papillae are asymmetrical, then hair will be curled.
Nobody really understands why you need a tightly packed-together mass of mesodermal cells in order to stimulate formation of hairs.
At certain locations around mouths of many species of mammals, special hairs are arranged in straight rows:
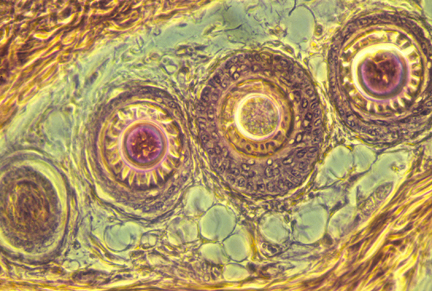


Bird feathers are arranged in very regular rows.

The regular spacing and alignment of feathers is caused by large papillae.
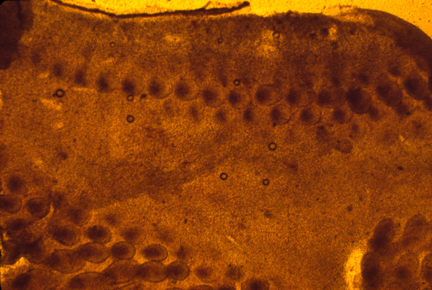
Feather papillae are often called "Feather Germs", for unimportant historical reasons.
Bird skin forms mesodermal papillae by active migration of cells into tight clumps, one clump at a time, row after row.
This is a good system for research on how regular anatomical patterns are formed. (not that we can be sure that they all use the same mechanism; but it is a big step forward to discover how any one spatial control mechanism works.) Maybe feather papillae are controlled by a "clock & wave-front system"? Or maybe by a "Turing system"?
The best research on spacing of bird feather papillae has been done in France. Scientists disagree whether the locations are first caused by changes in the ectoderm, which then cause mesoderm to form papillae; or whether the mesoderm aggregates first, and then the ectoderm responds.
One experimental method is to dissect the ectoderm away from the mesoderm, separating them before the patterns are formed, and observe which forms pattern first. Both results have been observed. So maybe it's a feedback loop?
Scales of snakes and lizards have even more exactly regular spatial arrangement, and are assumed to be controlled by mesodermal papillae.
One unexplained fact is that the regularity of spacing depends on whether the skin is stretched tight. If tension is reduced at the time of papilla formation, then papillae will form at irregular spatial locations.
Incidentally, this relation between physical tension and geometric regularity is a general phenomenon, even though it isn't mentioned in textbooks.
It was discovered primarily by Ruth Bellairs in London and Lev Beloussov in Moscow. A possible explanation is that physical forces are used to signal where embryological events should occur, so that changing the forces changes the patterns.
A good name for this is "The Bellairs'-Beloussov Phenomenon"
Neural Crest Ectoderm:
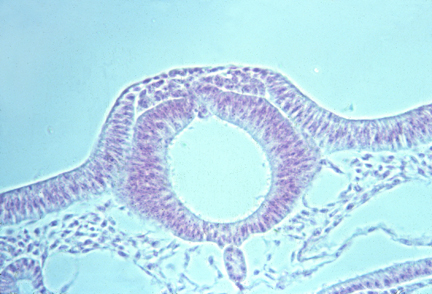
Development includes many cases of epithelial cell sheets folding and fusing their edges.
Many birth defects result from failure or incompleteness of fusion of epithelia:
Examples:
-
"Spina Bifida"
"Cleft Palate"
"Cleft Lip" "Hare Lip"
Retention of the oropharyngeal membrane includes a failure of fusion
Several heart defects result either from failures of fusion, or fusion at the wrong place. There are several versions of "Septal defects"
(Sometimes incomplete septum between left and right ventricles, (Sometimes this septum is on the wrong side of the aorta or the pulmonary artery
________________________________________________________________
What happens to the epithelial cells closest to where fusions occur?
In the neural tube, these boundary cells become mesenchymal and disperse.
In some placodes in the head, all their cells become mesenchymal and disperse.
In the lens placode and the otic placode, there is no conversion to mesenchymal cells.
In the fusion of the palatal shelves, cells undergo programmed cell death.
I don't know what happens at the fusion areas of the lip and face, or in the heart.
________________________________________________________________
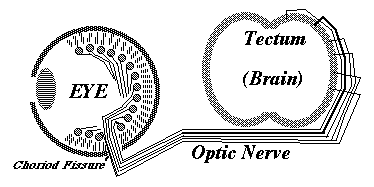
Retino-Tectal connection of the optic nerve.
The most-studied example, out of many (perhaps thousands) of examples of map-like connections of nerves from one area connecting to another area.
Sensory projections: skin to central nervous system Motor projections: muscles to central nervous system
Projections connecting one part of the central nervous system to another. (The retino-tectal projection can be regarded as brain-to-brain, because the retina forms as an outpocketing of the side wall of the brain.)

see page 96 in Scott Gilbert's textbook: And also pages 395-408

"Ephrin Ligands" and "Eph Receptors" are the names of two proteins found on the outside surfaces of some cells, including some nerve cells, and the cells that line arteries and veins.
There are at least two kinds of each, called type A and type B, with subtypes A1, A2, A5 etc.
They bind specifically to each other where cells touch; e.g. molecules of type A Ephrin Ligand on one cell bind specifically to molecules of type A Eph Receptors on any adjacent cell.
Both cells change in response to this binding. (Which is somewhat unusual, or at least hasn't been noticed or hypothesized much before. People tended to assume communications were just in one direction)
These molecules are used for chemotactic attraction and repulsion of cells, and for creating boundaries.
If a cell is stimulated to crawl toward whichever direction its surface has more ephrin binding, that is chemotactic "attraction" (although that is a behavioral response, not a physical force.)
Staining with fluorescent antibodies led to the discovery that embryonic retinas and parts of their brains have concentration gradients of eph receptors and ephrin ligands, in patterns that are not yet worth memorizing. At least six different selective adhesion proteins form gradients there.
Optic nerve fibers need guidance at several stages of their progress to, and through, the midbrain:
for example
-
"Attraction" toward the blind spot;
"Repulsion" away from the fovea;
Turning or not turning at the optic chiasma.
Projecting onto the tectum (in fish, amphibians & birds; but a different area in mammals (!))
People have been hoping to find such gradients for 40 years, and maybe they hoped too much, because simpler patterns would provide better explanations.
-
We shall not go unpunished,
Unless we keep it secret;
(Simplicity) of all things is what people need,
but complexity is more comprehensible to them.
-
Yeyo Svoye nujnaya lydyam; No slojnaya ponyatni im.
Meanwhile, I am trying to rewrite those Chemotaxis Pascal computer programs (that I projected in class, and that are posted on this course's web site) so that they can simulate formation of neural projections along concentration gradients.
Although it is not too difficult to invent behavioral properties that steer cells to the highest point in a concentration gradient, we need to invent behaviors that will cause them to find certain locations along concentration gradients, for example to go where the concentration is toward 40% maximum, while neighboring cells have behaviors that guide them, instead, toward where the gradient is 41% maximum. Not so easy; But I hope simple.
I think I have this figured out, based on Steve Roth's original idea. But I could sure use help, and appreciate suggestions and criticisms.