Fifth lecture Jan 27, 2016 Trying to catch up after two snow days
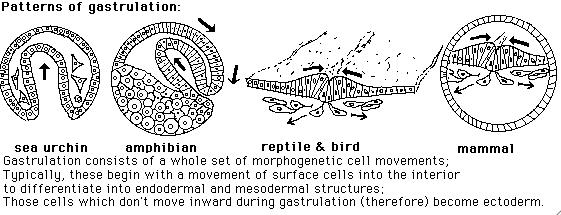
(I) Sea urchin embryos gastrulate by a combination of epithelial invagination and conversion of epithelial cells to mesenchymal cells ("ingression")
video shown in class Primary mesenchyme cells differentiate into skeleton cells (& secrete calcium carbonate rods)
Secondary mesenchyme cells differentiate into muscle cells
Skeleton and muscles are considered to be mesoderm.
The rest of the mesoderm, and all of the endoderm, develop from the
invaginated tube (called the Archenteron).
A smaller infolding near the animal pole (the stomodeum) folds inward and meets and fuses with the innermost tip of the archenteron.
-
Stomodeum -----> Mouth
Archenteron ----> Digestive tract
Blastopore ------> Anus
No neural tube forms in sea urchins.
Please notice that the blastopore does NOT connect to the blastocoel.
The blastopore connects to the archenteron.
The archenteron protrudes into the blastocoel.
"Trick questions" are often based on these facts.
--------------------------------------------------------------------------------
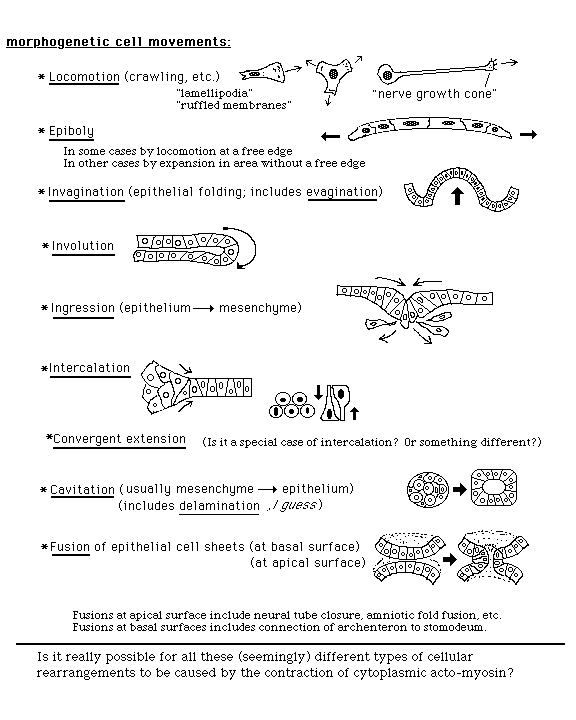
(II) Notice how often the geometric boundaries between different germ layers and future differentiated cell types depend on (or are controlled by?) invagination, ingression or other active cell movements.
Neurulation consists of folding half of the ectoderm into a hollow tube, and sealing the edges together. This process decides locations of boundaries between the following:
-
Neural Tube -----> Brain, spinal cord, (most) motor nerves, neural retina, pigmented retina.
Neural Crest = the two bands which fuse together --> Sensory Nerves, Pigment Cells, etc.
Somatic Ectoderm ---> Outer layer of the skin, lens of eye, inner ear, olfactory nerve
Folds become boundaries between differentiated cell types.
Other examples occur in the formation of the eye, gill slits, & inner ear.
There are many other examples, in all kinds of embryos.
(The cause isn't yet understood: and the popular theory of "positional information"ignores such creation of boundaries by folding etc. Instead, differentiation is claimed to be caused by chemical diffusion gradients,
and gradients definitely do occur in developing embryos, and can control pattern formation)
--------------------------------------------------------------------------------
(III) Frog and Salamander embryos gastrulate by "Involution" (which is a variation of invagination in which cells sort of roll around the edge of the blastopore.
"Evagination" is an abnormal movement of cells outward, leaving the ectoderm as an empty sack, connected to an inside out blob of what would have been mesoderm and endoderm.
Probably, involution is caused by a wave of acto-myosin contraction, but more knowledge is needed.
Personally, I spent years trying to map physical forces inside gastrulating salamander embryos, but I failed.
If I have time, I will tell you the analogy between involution and some luggage conveyer belts at airports.
Amphibians (and all vertebrates) form a stomodeum, which connects to the archenteron, & forms the mouth & throat.
Mouth structures (teeth, pituitary, some salivary glands) are (somewhat arbitrarily) considered ectodermal.
--------------------------------------------------------------------------------
(IV) Gastrulation in birds, mammals and reptiles occurs by ingression (conversion of epithelial cells to mesenchymal cells) anterior-posterior along a thickening called the primitive streak.
The anterior end of the primitive streak is called Hensen's Node (synonymous with primitive node). Cells that ingress at Hensen's node differentiate into the notochord. (a solid cylinder of vacuolated cells tightly wrapped by helical fibers of type I collagen. That is the same protein that tendons & fascia are made of.)
The rest of the mesoderm and most of the endoderm develop from cells that ingress along the rest of the primitive streak.
Bird and reptile embryos gastrulate on flat sheets of epithelial cells that are stretched on part of the surface of "the yolk" (in the sense of that yellow sphere, frequenty seen at breakfast).
Mammal embryos gastrulate like birds (because we are both descended from reptiles)
Mammal primitive streaks form on flat sheets of cells stretched across one end of a hollow ball of cells, called the blastocyst.
The sheet of cells is part of the "inner cell mass"
--------------------------------------------------------------------------------
(V) Three different ways that identical twinning occurs in mammals.
-
Two primitive streaks form, side by side, on the same sheet of cells. (< 1%)
Two inner cell masses form inside the same blastocyst. ~ 49%
Two blastocysts form by rearrangement of cells from one embryo. ~ 49%
It is interesting that people guess so confidently about human embryos.
Conjoined twins are a small percentage of the twins that develop side by side in the same sheet of inner cell mass cells.
Questions for class discussion:
Guess how you can know which of the three causes of twinning has occurred in any given case.
(Hint: based on something about the placenta)
Guess why conjoined twins are (almost) always mirror images of each other.
(e.g. joined hip to hip, belly to belly, back to back, or head to head, etc. etc.
but never back to head, or anything to something different)
Exceptions occur when one twin is much larger than the other.
Conjoined twins are never joined arm to arm or leg to leg, much less arm to leg or arm to back.
People sometimes regard conjoined twins as originally separate, and then fused together.
Another popular guess is that what had been one embryo has become split apart.
Alternatively, visualize two sculptors, both carving the same piece of rock or clay.
Does the mirror-image rule support or disprove any of these possible causes for conjoining twins?
--------------------------------------------------------------------------------
(VI) In the late 1800s, the words gene and genetics had not yet been invented, but researchers realized that something in chromosomes must cause cell differentiation in embryos.
They guessed that different genes probably got distributed to different parts of each embryo; for example, skin cells might get only the genes needed to make skin, nerve cells might get only genes for being nerve.
This guess was mistaken: All the differentiated cells of the body contain the same set of chromosomes and genes. Differentiation works by "turning on" only a subset of genes (where "turning on" means stimulating transcription and processing messenger RNA for a given gene).
Instead of skin only getting skin genes; only skin cells "turn on" skin genes, and so on for all the 250 differentiated cell types.
A few kinds of animals selectively destroy and "throw away" certain chromosomes in all cells except those which will differentiate into sperm and oocytes. Discovery of this throwing away phenomenon either stimulated or seemed to confirm the mistaken idea that differentiation works by handing out different genes and/or different chromosomes to cells in different parts of each embryo.
This throwing away phenomenon is called "chromatin diminution" but you don't need to memorize that.
Wilhelm Roux did a famous experiment intended to confirm the theory of different parts of each embryo getting only certain genes. He poked red-hot needles into 4 or the first 8 cells or early frog embryos; or sometimes poked hot needles into 2 of the first 4 cells, or 1 of the first 2 cells.
The idea was to kill or severely damage half the cells of an early embryo, with the expectation that the undamaged cells would continue and develop into the same tissues that they normally form, even though the "killed" cells failed to differentiate.
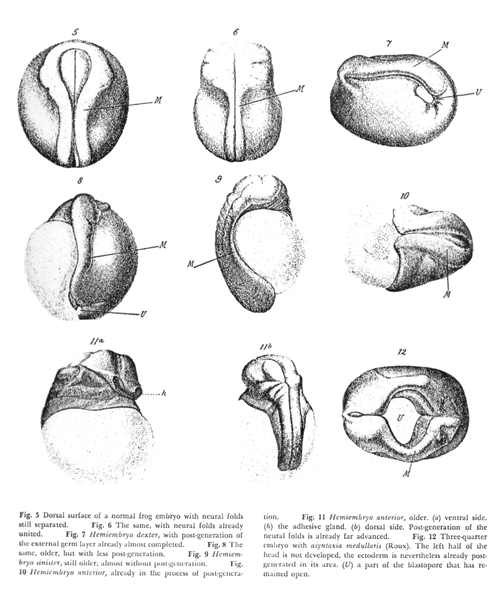
Textbooks use the verb "killed" to refer to the effect of the hot needle on embryonic frog cells. Actually, the original paper says the "killed" cells were still alive, and that after several hours they began to divide again.
Anyway, people interpreted the result as support for the idea of different genes being distributed unequally to different cells. (Nobody really understands the results of this experiment, but it has been repeated with the same results)
Ten years later, Hans Driesch, tried another experiment to test the same theory. He shook water that had 2 and 4 cell starfish embryos in it. Some of their cells got separated apart. He expected they should develop into half embryos or quarter embryos.
The results astounded Driesch (and everybody else).
When the first 2 cells separated, each developed into a half-sized, normally proportioned pluteus. If one of the first 4 cells were separated, it developed into a quarter-sized, but normally proportioned pluteus, with each part a quarter normal size.
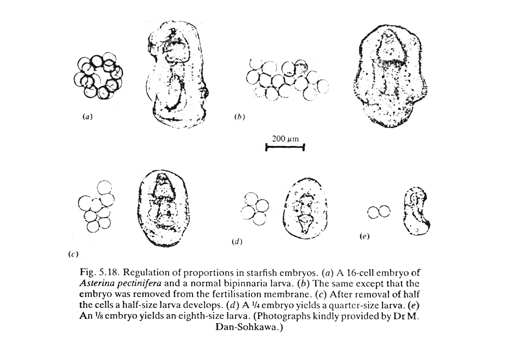
This disproved the theory of different parts of embryos receiving different genes.
But even today scientists don't really understand the mechanism that changes the sizes of parts of embryos in proportion to the size of the whole embryo.
--------------------------------------------------------------------------------
Study guide: Make sure that you are familiar with the meaning of all the following terms, and their significance for embryology.
invagination
ingression
primary mesenchyme
secondary mesenchyme
archenteron
stomodeum
blastopore (does NOT connect to the) blastocoel
neural tube
neural crest
somatic ectoderm
"theory of positional information"
Roux
Driesch
Be prepared to answer the following questions:
a) The 3 different ways that identical twinning occurs in mammals.
b) If it were really true that twins become conjoined because of being pushed together, then could non-identical twins sometimes become conjoined, in addition to identical twins?
c) Sometimes, one of a pair of identical twins has all his or her heart, stomach and liver arranged and shaped so that they are mirror images of normal hearts, etc. (e.g. The aorta comes out the right side of the heart and the pulmonary artery comes out the left side.)
Suggest a hypothesis to explain how and why this happens.
d) What might happen if you pushed two starfish or sea urchin embryos tightly together side by side?
(Answer: it develops into a double-sized pluteus)
How is this experiment related to what Driesch did?
e) What might happen if you killed an embryo of a sea urchin or a frog embryo at the one celled stage,
and pushed this dead cell tightly against the side of an undamaged embryo at its one cell stage?
Hint: Suppose that the results were analogous to what happened in Roux' experiment.