Evolutionary Developmental Biology, the so-called "Evo-Devo"
Embryological phenomena have different kinds of causes:
- Functional
- Genetic
- Mechanical
- Evolutionary
Historically, scientists have concentrated first on one kind of cause, then focused more on some another kind, then switched to another.
Ernst Haeckel's ideas were dominant between about 1870-1900 or later
He invented a famous cliché: "Ontogeny recapitulates phylogeny."
Meaning that the shape changes of embryonic development are speeded-up versions of changes that occurred during evolutionary history of each kind of animal.
A concept based on mis-application of Darwin's evolution.
Recapitulation is an exaggeration; but with some truth.
Gill slits, notochord, very similar cell lineages among spirally-cleaving species: are all explainable only by shared ancestry.
If we hadn't evolved from fish, would our embryos have gill slits?
Even though we now use our gill slits to form the ear canal, the tonsils, the thymus "gland", and the parathyroid gland.
Notochords are evidence that both we and fish, and also sea squirts, have common ancestors. (Also remember Spemann and the inductive effect of the notochord.)
*********************************************************
In the last couple of decades there have been big surprises about certain families of transcription factor genes that control organ formation.
For example, insects have compound eyes, with a surface retina facing outward.
Vertebrates have camera eyes, with the retina on the inside, with light focused on it by one lens
Yet some of the transcription factors that control the locations of eye development have very similar amino acid sequences in flies as in mice, e.g. the Pax6 gene
Putting copies of mouse genes and Pax-6 proteins into flies causes them to form compound eyes in abnormal locations.
Putting copies of fly Pax-6 genes into Xenopus frog embryos can cause them to form extra eyes at abnormal places.
But fly tissues form compound eyes, and vertebrate tissues form "camera eyes" (hollow, with a retina & lens)
Maybe both kinds of eyes evolved from some very simple eyes or eye-spots of flatworms?
And the transcription factor proteins that stimulate expression of the rest of the eye genes stayed the same, cven though vertebrates then evolved completely different kinds of eye structures than arthropod eyes? It's anybody's guess. What do you think?
++++++++++++++++++++++++++++++++++++++++++++++++++++++
Hearts have very different locations and structures in arthropods as compared with vertebrates (almost the reverse geometry, upside-down
Heart in the chest in vertebrates: neural tube runs down the back.
Arthropod neural tube encircles the throat, and runs rearward down the belly.
But tinman genes were found both in flies and in vertebrates
And very similar distal-less genes are expressed (locally transcribed)
- At the outer ends of fly legs
- Along the outer edge of vertebrate limb buds
- At the ends of sea urchin tube feet
- In the middle of spots on butterfly wings?
Hox gene expression patterns:
Genes nearer the 3' end of sets of hox genes are expressed nearer the head of the embryo.
"The colinearity phenomenon" (nobody understands the mechanism yet).
Deletion of individual hox genes can cause one segmental structure to be replaced by an extra copy of some structure that is normally next to it.
This is analogous to replacement of your thumbs by an extra index finger!
(Which does NOT actually happen! I just invented the possibility to dramatize the point.) And I don't know why vertebrates don't replace thumbs if Hox d-9 were deleted.
A real example is formation of cervical vertebrae by thoracic-like vertebrae with ribs ("Cervical ribs", in your neck, can be very painful)
[If you're interested in learning more about hox genes, the conserved homeobox region in the proteins they encode, etc., see this Wikipedia article.]
===========================================================
SOME MAJOR DIFFERENCES IN INSECT EMBRYOLOGY (among which fly development is somewhat special)
1) All 3 axes (anterior-posterior, dorso-ventral, medio-lateral) are already irreversibly determined before fertilization by signals and mRNA from surrounding maternal tissue.
Notice the contrast to vertebrates, in which axes get decided, one by one, later in development
What should we conclude when the Wnt gene turns out to be part of how frog eggs react to sperm entry or gravity in deciding which end should form the head?
One idea is to think of Wnt, etc. as analogous to transistors and capacitors etc. that you can find in TVs and air-conditioners and computers etc.
2) Arthropod embryos don't cleave into separate cells until the 13th mitotic cycle (~6-thousand cell stage). Until then they are syncytial, so that even fairly large proteins like transcription factors are free to diffuse around.
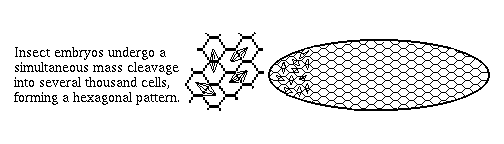
This is very different from vertebrates, and the great majority of phyla, in which nuclei are put in separate cells soon after fertilization, and before many decisions are made.
Even small proteins can't diffuse from cell to cell.
3) In insect eggs, most of the oocyte cytoplasm is produced in 15 mitotic sister cells, called nurse cells. These are not at all the same as follicle cells, although follicle cells perform a nursing function, and insects have follicle cells in addition to nurse cells. Vertebrates have follicle cells, but not nurse cells.
4) In insect eggs, mesoderm is internalized along the bottom, in an invagination process that looks like neurulation in vertebrates.
In insect eggs, endoderm folds inward from both ends toward the middle, in a process separate from the internalization of mesoderm.
In insect eggs, neural ectoderm is internalized by many separate cells, ingression on the lower surface.
5) In insect eggs, the ventral side is the center of embryological movements and differentiation, instead of the dorsal side as in vertebrates.
There is a very old theory, now supported by some new molecular evidence, that arthropods evolved as upside-down equivalents of vertebrates, or that vertebrates evolved upside-down, relative to arthropods. Heart on the top; Nerve cord on the bottom! This old theory turns out to be supported by discoveries of in what places several transcription factors are synthesized.
Some important Evo-Devo research that was done here in the UNC Biology Department:
Prof. Anne Burke and her Ph.D. student, Julie Nowicki (who was a Teaching Assistant in this course for two years) collaborated with Prof. Alan Feduccia to test
whether the 4 toes of bird feet correspond to toes 1, 2, 3, & 4,
instead of toes 2, 3, 4 & 5,
based on which D hox genes are transcribed in the toes of ostrich embryos.
This tested which reptiles birds evolved from (whether from dinosaurs, or other groups).
Their evidence contradicted the most popular "birds are dinosaurs" theories.