Lecture Notes for Wednesday, January 31
"Embryonic Regulation"
Hans Driesch: discovery of "Regulative development" (in 1890s) He separated the first 2, and the first 4 cells of starfish embryos, and observed their development into two half-size plutei, or 4 quarter-size Conversely, you can push two one-cell stages together and sometimes they will develop into double-sized plutei.
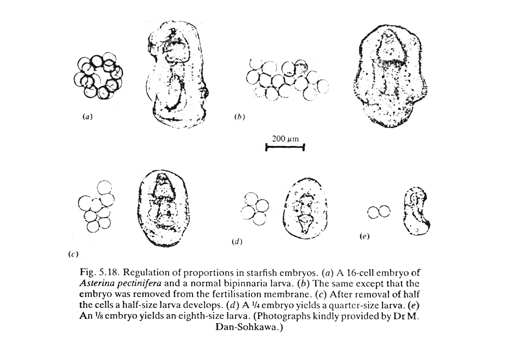
This disproved the theory of different parts of embryos receiving different genes.
But even today scientists don't really understand the mechanism that changes the sizes of parts of embryos in proportion to the size of the whole embryo.

But please don't be misled by the word "regulation",
which can seem to imply some exterior control telling somebody what to do.
This kind of regulation is internal adjustment, to fix damage or overcome abnormality or effects of some external disturbance.
Whatever adjustments cause regulation of shape and adjustment of proportionality of sizes, sometimes require time to adjust.
"Regulative" development contrasts with "mosaic" development, in which cell fates are pre-determined, for example in the nematode Caenorhabditis elegans.
Driesch concluded that embryos are somewhat supernatural
in the sense of being controlled by something mind-like.
An even better example of size adjustment of parts in proportion to wholes is Dictyostelium formation of fruiting bodies, stalks, and length/width ratios of slime mold slugs.
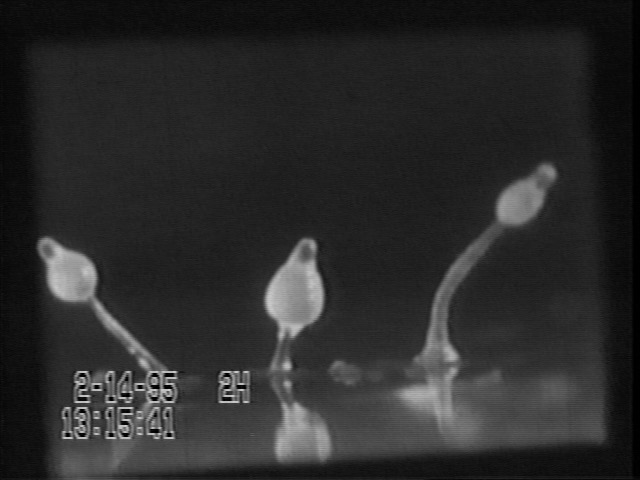
(Please understand that these "slugs" are masses of a kind of amoebae, & only happen to resemble real slugs, which are a kind of snail.)
Dictyostelium discoideum is the most-studied one of the "Cellular Slime Molds". In one stage of their life cycle, a culture consists of small amoeboid cells that move around, eating bacteria. When they run out of bacteria, they aggregate by chemotaxis, and form multicellular "slugs" that can crawl around. The slugs vary widely in size, depending on how many individual amoebae aggregated to form them. They then differentiate into stationary "fruiting bodies" that consist of a tapering stalk, with a lemon-shaped mass of spores at the top, and have the same proportions over a size range of at least 500.
This is even more than Driesch's revolutionary discovery that starfish embryos can "scale" over a sixteen-fold size range. (Incidentally, this most-studied species was discovered by a UNC undergraduate [Kenneth Raper] on some horse dung in Duke Forest) For many years, the leading researcher on slime molds was Lindsay Olive, who was a professor in this department, and kindly provided many different species to use in laboratories of this course.
You can cut a Dictyostelium "slug" into 10 or 100 pieces, and the cells of each fragment will reorganize to form a normally-proportioned slug, and then a scale model stalk and spore mass, one one-hundredth the size the original "slug" would have formed. Echinoderm plutei can develop "scale models" over a volume ratio of 8 to 1 or sometimes 16 to one.
Scaling may have different mechanism in plutei than in slime mold fruiting bodies.
But I regard this as one of the 3 or 4 deepest questions in biology.
What does the ability "to scale" tell us about normal mechanisms that cause
organ formation?
That's the big question. There might be several answers, or one.

Lewis Wolpert's hypothesis of sloping diffusion gradients combined with quantitative thresholds for induction of alternative cell types.
He calls this the French Flag model, because it can produce scale-model French flags:
1/3 red, 1/3 white, 1/3 blue.
If you google "French flag embryo" images you can see many relevant illustrations.



Flags generated and then distorted by a computer program


Wolpert's French Flag model applied to flatworms

The caption on this is from a lecture given many years ago, and is not a potential exam question. However, you might want to think about it.
Wolpert's model is generally assumed to mean gradients of a diffusible substance. However, gradients can also be created without anything moving, by adjusting properties to conform with neighboring locations.
"Diffusion" effect created by a cellular automata program that computes the numerical value of nearest neighbors.
-----------------------
Roux "hot needle" experiment (~1880s)
Wilhelm Roux did a famous experiment intended to confirm the theory of different parts of each embryo getting only certain genes. He poked red-hot needles into 4 or the first 8 cells or early frog embryos; or sometimes poked hot needles into 2 of the first 4 cells, or 1 of the first 2 cells.The idea was to kill or severely damage half the cells of an early embryo, with the expectation that the undamaged cells would continue and develop into the same tissues that they normally form, even though the "killed" cells failed to differentiate.
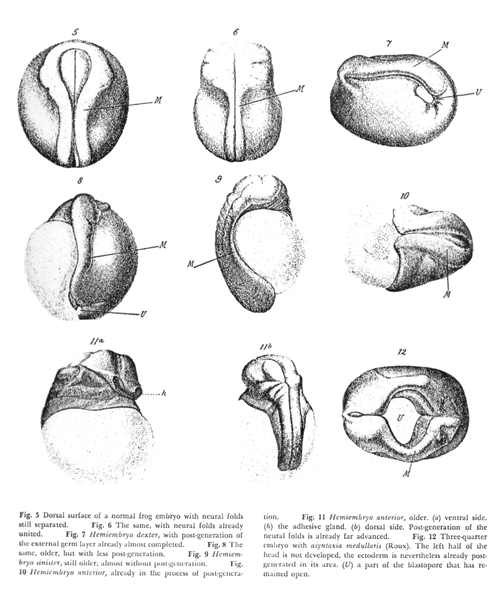
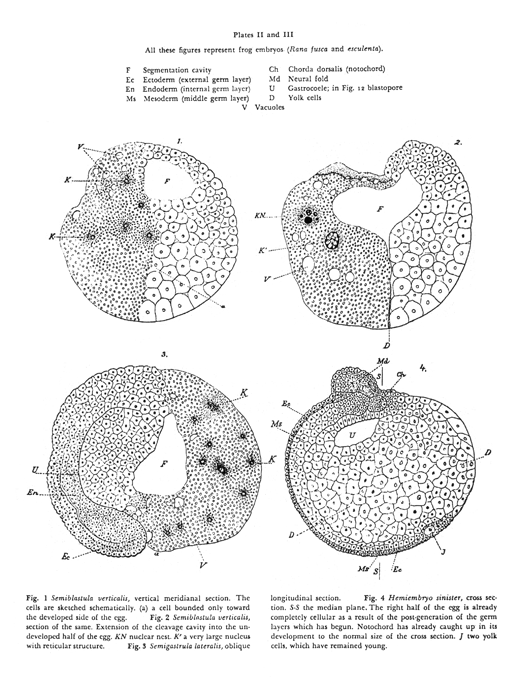
Textbooks use the verb "killed" to refer to the effect of the hot needle on embryonic frog cells. Actually, the original paper says the "killed" cells were still alive, and that after several hours they began to divide again.
Anyway, people interpreted the result as support for the idea of different genes being distributed unequally to different cells. (Nobody really understands the results of this experiment, but it has been repeated with the same results)