Lecture notes for Monday, January 29, 2018
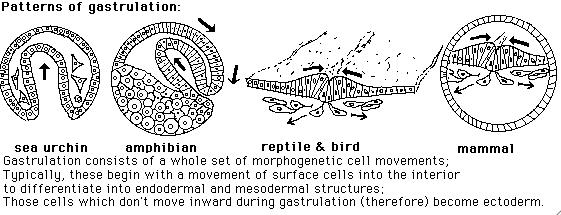
Mammal embryos cleave like sea urchins (because mammal eggs have little yolk) but gastrulate like birds (because mammals evolved from reptiles, with lots of yolk)
Meroblastic cleavage: Teleost fish Birds (& Reptiles) Holoblastic cleavage: Sea Urchins, Salamanders and Frogs, Mammals The more "yolky" the oocyte, the more meroblastic the cleavage The less yolky the oocyte, the more holoblastic the cleavage. Gar Fish, Bowfins & Sturgeon have holoblastic cleavage (and are NOT teleosts)
Some frogs and salamanders that lay eggs on land have meroblastic cleavage Three species of mammals lay yolky, shelled eggs, with meroblastic cleavage.
Less yolky embryos gastrulate more by invagination or involution. Hensen's node is the place on the primitive streak where ingression begins. It is analogous to the dorsal lip of the blastopore in amphibians. Mammals reverted to holoblastic cleavage but continue to gastrulate like reptiles (which means: like birds). Something non-typical and extreme about teleosts is progressive formation of body organs, from head to tail. Bird embryos are somewhat this way, but most vertebrates aren't.
The early mammalian embryo is a hollow ball of cells called the blastocyst. Only a small fraction of the cells will become part of the embryo. The rest form membranes and the support structure for the developing embryo.
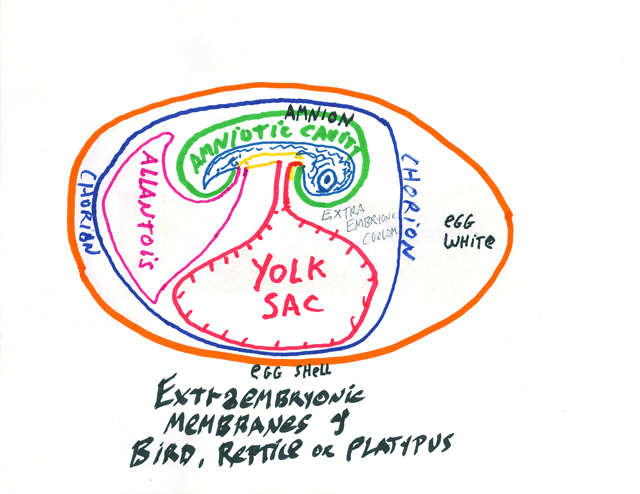
Reptiles evolved four extra-embryonic membranes: Chorion: Ectoderm & Mesoderm
Is in direct contact with the uterus (in mammals) [of course, mammals are the only vertebrates that have uteruses]
Amnion: Ectoderm & Mesoderm The space enclosed inside the amnion is the "Bag of Waters".
Yolk sac: Endoderm & Mesoderm Allantois: Endoderm & Mesoderm is an outpocketing of the hindmost part of the digestive tract (Stores urine) (very well-developed blood vessels)
Birds and mammals inherited all 4 of these extra-embryonic membranes.
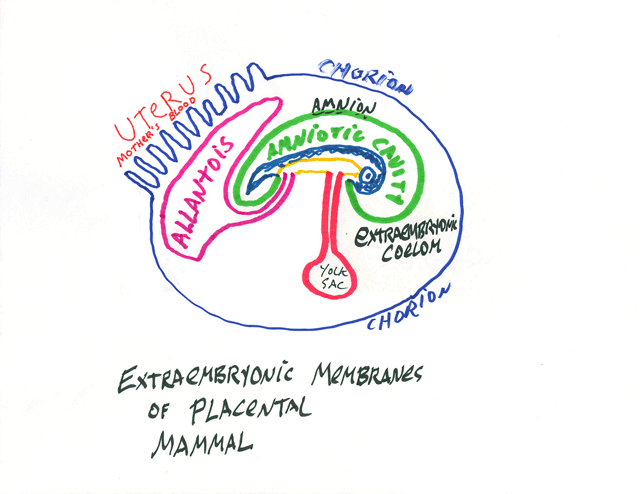
The umbilical cord connects all four of these to the body itself,
and contains two arteries and one vein. Turtles, snakes, lizards sometimes hatch with a large yolk sac protruding from them. This will be physically retracted into the body in a few days; don't cut it off! Many species of snakes incubate their eggs inside the uterus, instead of laying eggs. Most reptiles have flexible "leathery" egg shells. Indentations in reptile egg shells often push on early embryos, which can result in two-headed offspring. In mammal embryos, the body develops out of part of the inner cell mass. The large sphere of cells ("the trophoblast") develops into the chorion. Mammal embryos digest a pathway into the wall of the uterus "endometrium" and crawl down into it until completely surrounded by uterine cells. (This digesting & burrowing is called "implantation")
digest through uterine tissues, including blood vessels,
so that blood flows directly across the part of the chorion where the allantois is just inside.
Pigs, horses and many other kinds of mammals have much less of this tissue removal.
Note: There is no kind of animal in which the circulatory system of the mother ever connects directly to the circulatory system of embryos (like a trailer hook-up).
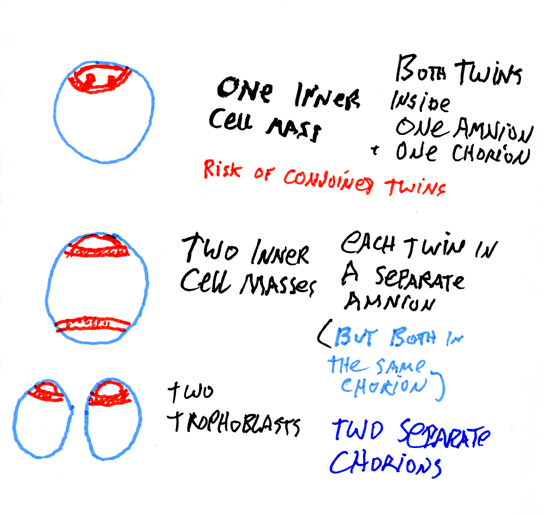
Three alternative causes of identical twinning:
If both twins are enclosed in the same amnion, then there were 2 primitive streaks in the same inner cell mass (and they will also be inside the same chorion). This happens in about 1% of identical twins. This is the only case in which formation of conjoined twins is possible.
If both twins are together inside the same chorion, but each separate in their own two amnions, then they developed from two separate inner cell masses, inside the same trophoblast. About half of identical twins develop this way.
If twins are both in their own separate chorions, then they developed from separate trophoblasts, and might not be identical twins.
Incidentally a very great immunological discovery was accidentally made by using skin grafts to (try to!) distinguish between cattle twins that were genetically identical versus which were "fraternal" twins.
Researchers expected that only non-identical twins would "reject" organ grafts from each other.
But immune tolerance VERY unexpectedly developed in non-identical twin cattle.
Their immune systems didn't attack each other! Can you figure out, or guess, why not?
(Note: this doesn't occur in human twins, for some unimportant reason.)
************************************************************************
Induction of neural tubes by notochord mesoderm: "Primary Induction"
Other examples include induction of the lens by the eye cup,
Induction of salivary gland epithelia by salivary mesenchyme,
Induction of frog mouth suckers by salamander facial mesenchyme
Chordin is one of several proteins that have been discovered to
induce neural tube, etc.
Two obstacles to discovering the molecular signals that produce induction
Prof. Bob Goldstein of the UNC Biology Dept discovered the first example
of induction in Nematode embryonic development.
This was an extremely important discovery, because unexpected by many.
Many expected induction to occur in species with "regulative" development.
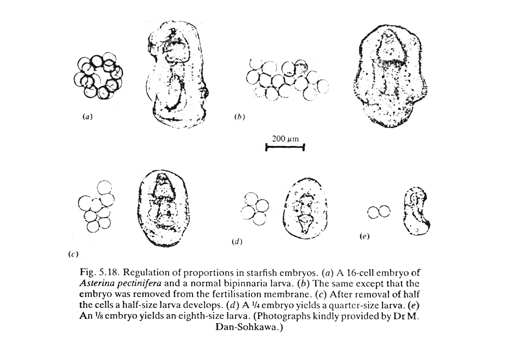
Hans Driesch: discovery of "Regulative development" (in 1890s)
He separated the first 2, and the first 4 cells of starfish embryos,
and observed their development into two half-size plutei, or 4 quarter-size.
Conversely, you can push two one-cell stages together and
sometimes they will develop into double-sized plutei.
This disproved the theory of different parts of embryos receiving different genes.
But even today scientists don't really understand the mechanism that changes the sizes of parts of embryos in proportion to the size of the whole embryo.
|