Embryology Biology 441 Spring 2011 Albert Harris
Lecture notes for Monday, Feb 7, 2011

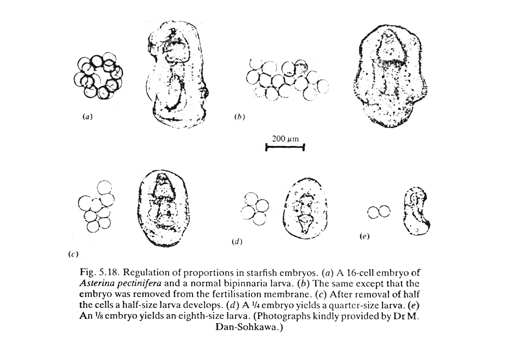
Hans Driesch (1890s), doing research at marine laboratories in Trieste and Naples, (originally using early embryos of starfish), discovered:
"Embryonic Regulation"
But please don't be misled by the word "regulation",which can seem to imply some exterior control telling somebody what to do.
This kind of regulation is internal adjustment, to fix damage or overcome abnormality or effects of some external disturbance.
Embryos of many kinds of animals are not regulative.
They are said to have "mosaic development"
More regulative development <-------------> More mosaic development:Mammals Echinoderms Amphibians Sea Squirts Flies Snails Nematodes
-
If species use ooplasmic segregation
their embryos development will be mosaic.
Very consistent cleavage patterns,
and consistent cell lineages
occur in species with mosaic development.
(Because the mechanistic causes are not yet understood well enough.)
Many people think the key difference is just how soon in development
cells become irreversibly committed to differentiate into specific cell types,
and to form particular parts of the body.
The sooner cells become irreversibly committed,
The more mosaic the development of those kind of embryos will be.
Other people think that some fundamental differences in mechanisms are the cause of development being more regulative or more mosaic.
(for example if differentiation were controlled by special proteins or other molecules being put into particular cells at the time of cleavage, that would cause development to be more mosaic.)
Regulative development would be impossible without cell-cell communication. (I guess?)
The theory called "Positional Information" claims that regulative development proves that cell differentiation is controlled by diffusion gradients of "morphogens", the local concentrations of which control which differentiated cell type each embryonic cell will differentiate into.

Driesch, himself, concluded from regulation that the mechanisms that control embryonic development must be in some sense supernatural.
He used the word "entelechy" to mean the controlling consciousness.
(My PhD. Major Professor, J. P. Trinkaus used to joke that embryos would need at least TWO entelechies, one for each separated cell.)
If you are willing to include cellular slime mold amoebae, and their formation of stalked "fruiting bodies" as an example of something like embryos, then their development is the most regulative of all, because you can cut a Dictyostelium "slug" into 10 or 100 pieces, and the cells of each fragment will reorganize to form a normally-proportioned slug, and then a scale model stalk and spore mass, one one-hundredth the size the original "slug" would have formed.
(Please understand that these "slugs" are masses of a kind of amoebae, & only happen to resemble real slugs, which are a kind of snail.)
About ten years before Driesch's discovery of embryonic regulation, another great German embryologist named Wilhelm Roux tested the effect of 'killing' one of the first two cells of amphibian embryos at the two-cell stage. (He also tried killing two of the first 4 cells, and different combinations of 4 of the first 8 cells.)
(He 'killed' these cells by poking them with red-hot needles! And if you read Roux's actual paper, you find out that the cells Were not quite dead, and sometimes started dividing and developing later.)
Roux's results were very different from Driesch's.
His unkilled cells developed into half-embryos,
in the sense that only half the organs and structures formed.
Roux's famous "hot needle" experiment has been successfully repeated by others,
Including in some other kinds of animals (including sea squirts)
And had led everyone to believe that embryonic development works by
each dividing cell being given different information (perhaps different genes)
at the time of each mitotic division.
Driesch's discovery of "Embryonic Regulation" conclusively disproved
Roux's theory. But nobody knows (and nobody even ASKS!)
why "killed" damaged cells prevent embryonic regulation.
No theory explaining Driesch's discovery can be true unless it also
can explain Roux's observations.
Regulation is one of several embryological phenomena in which the same end results are reached by any of two or more series of intermediates.
Another example is "secondary neurulation".
(The neural tube can be formed either by epithelial folding,
or formed by cavitation of a solid mass of cells)


The most extreme example is sorting out of separated embryonic cells
to form functional, or sometimes even normal anatomy.
(Cell sorting will be discussed in a later lecture).
My own theory has been that all such regulation phenomena
Should be regarded as logically equivalent to homeostasis
(as in physiological homeostatic control of body temperature)
with the difference that geometric properties are being regulated
instead of simple quantitative variables.)


Homeostatic control depends on stable balances of opposing forces.

Many people think that stable balances between opposing forces is the same thing as minimization of thermodynamic free energy or minimization of potential energy.
This is a common mistake.
Some forces are "conservative" in the sense that they do not continue to expend energy in a situation of static counter-balance.
Other forces are "non-conservative" in the sense that the use up energy even when not producing any change, and exactly counter-balanced by some other force.
Not understanding this leads people to make two further mistakes:
-
1) They think that stable counter-balances are the same thing
as minimization of free energy.
2) They think that only conservative forces can cause stable counter-balance.
(I am sort of a fanatic on this subject, because I think these fallacies hold back embryology, including accurate understanding of cancer.)
Stable counterbalances of both non-conservative forces (like contraction of acto-myosin) and conservative forces can explain shape and size control of many structures and organs.


A big problem is how such mechanisms adjust sizes of parts in proportion
to sizes of wholes, which is closely related to adjusting sizes
(and relative strengths of forces) in perpendicular directions.
(which means keeping the same shape at different sizes)
This sometimes occurs in non-living mechanical patterns, from which we can learn important lessons.


When a separated embryonic cell rounds up, few people have trouble this rounding is directly caused by mechanical forces. But the later adjustment of fates of cells is not thought of as having anything to do with mechanical forces.
Notice, however, that in Roux's experiment the continued presence
of the "killed" cells prevents the remaining undamaged parts of
the embryo from rounding up.
So maybe anything that prevents reshaping would block regulation?
There are lots of important phenomena left for you to discover.
(But I warn you that frogs tend to lay eggs in the middle of the night
on weekends, so it's not so easy to get their eggs at the 2 or 4 cell stages!
So, it's not so easy to repeat variations of Roux's hot needle experiment.)
One way to avoid having to explain embryonic regulation is to concentrate research on the kind of animals whose development is more mosaic than any others'! (Specifically, nematodes)
Possible topics for discussion:
Do Nematodes not have entelechies?
What if Driesch had done his experiments on embryos of a species that has mosaic development?
(Roux' false conclusions about unequal distribution of genetic material would have seemed to be confirmed?)
The heart develops from lateral plate mesoderm, at its extreme lateral edge where the left and right lateral plates come together. If a barrier is put between them, so that they cannot touch, then each develops into an entire heart. Two hearts! Another example of embryonic regulation.
Other examples are that branched legs will form if you split limb buds, that double eyes will develop if you split the optic cup, and that single cyclops eyes will develop (only 1) if you put the optic cups too close together.







