Embryology Biology 441 Spring 2011 Albert Harris
Cell locomotion, traction, and sorting
Malcolm Steinberg's "thermodynamic theory", Cadherins, "Haptotaxis", "Traction" and all that..
And also some class discussion, I hope, about whether theories help or hurt the progress of research, and reasons why or why not.
Most of the cells of the body can crawl, especially cells in embryos.
(They crawl like amoebae, especially like Dictyostelium amoebae)
Body cells propel themselves by exerting"traction" force tangentially through certain areas of their plasma membrane.
This was discovered by putting tissue culture cells on thin gels (with marker particles embedded in them), and later by putting cells on very thin sheets of silicone rubber, which is visibly wrinkled by traction forces.

Body cell locomotion is NOT caused by cycles of protrusion, adhesion and contraction (with the possible exception of nerves), but many textbooks still say that is how they move.
Because of a theory that biochemical effects cause gels to shrink
many scientists were not convinced by the distortion of gels.
Silicone rubber settled the issue, because it is inert,
and because wrinkles are visible in still photographs.
Yu Li Wang and Micah Dembo use acrylamide gels & track movements
of embedded plastic beads, using special computer programs
to calculate the locations, directions and strengths of cell traction.
They call this "Traction Force Microscopy" "TFM"
(they are in Boston) Other elastic substrata have been developed.
Many differentiated cell types exert strong enough traction to move other materials around & arrange them in anatomical patterns.
For example, sponge cells move spicules (pieces of skeleton) And vertebrate cells move and align collagen (the protein that tendons, & many other structures are made from) [sponge cells also rearrange sponge collagen = "spongin"].
According to one theory, all ligaments, tendons, muscles, and
part of the walls of arteries are generated by mesenchyme cell traction.
Some of the evidence is that cell traction will form artificial tendons
and muscles in tissue culture, and that collagen can be purified from
rat tail tendons, then covalently labeled with fluorescein,
and when injected into chicken embryos, this labeled collagen
gets rearranged to form tendons, artery walls and other normal structures.
These fluorescent collagen injection experiments were devised to disprove competing hypotheses that collagen is secreted in place or that it self-assembles in response to local variations in chemical environment.
These experiments were done by David Stopak at Stanford University, after he did his PhD in the UNC Biology Department, and was a TA in this course.
Crawling tissue cells can be guided and shaped by adhesion gradients and spots. (which can be made by heat evaporation of gold and other metals in a vacuum). This was discovered by an English MD & pharmaceutical researcher named Stephen Carter, who named the phenomenon "haptotaxis"
[70 years earlier, the German embryologist Ernst Haeckel had guessed that body cell movements might be guided by adhesion gradients, and independently invented a name for this. "Haptotaxis" ]
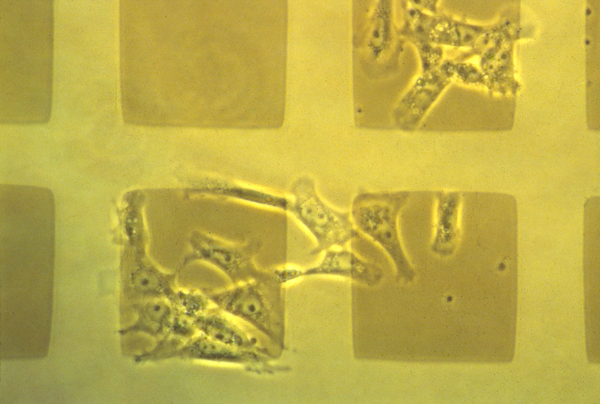
Haptotactic accumulation of cells on the most adhesive surface available occurs by a sequence of many tug-of-war events, in which the stickier end "wins".
In the following photograph, mouse cancer cells were given 4 choices:
-
a) Uncoated glass
b) Metal coated glass
c) Metal coated cellulose acetate
d) Uncoated cellulose acetate

Both cancerous and non-cancerous cells were cultured on grid patterns that gave cells a choice which of these 4 surfaces to accumulate on:
The result was:
Cells accumulate most a bc d Cells avoid most
Which can be the explanation?
-
Phototaxis? Some respose to electrical conductivity of metal?
Relative strengths of cell-surface adhesions?
How much adhesion proteins (fibronectin) absorb to different materials?
(& are very small cells, despite their name)
preferentially on these grid patterns in the exact opposite order of preference
d cb a
Prof. Abby Rich, formerly at NYU Medical School, did excellent research on this; but it is still a major puzzle, with potential medical implications.
Can you invent alternative hypotheses that might be able to explain these phenomena?
Can you invent experiments, except by first inventing theories for the experiments to test?
8 important researchers on cell rearrangement mechanisms and functions
Henry Van Peter Wilson at UNC from 1899 to his death in 1937 (still doing research); see portrait below
Hans Holtfreter, PhD Student of Spemann; fled Hitler to Canada & Rochester N.Y.
Malcolm Steinberg at Johns Hopkins, then Princeton; very recently retired
John Philip Trinkaus. At Yale (recently died) [Great-Nephew of John Philip Sousa, the composer]
Albert Harris At UNC since 1972 Previously student of Trinkaus and Abercrombie
Calhoun Bond, at UNC, T.A. in this course, now a Professor at Greensboro College; bottom right in the picture of Wilson's portrait, below
M. Takiechi In Japan; main discover of cadherins, which are a family of qualitatively specific cell-cell adhesion molecules. (qualitative difference = difference in kind)
Rachel Fink Professor at Mount Holyoke College; previously grad. student at Duke
Discovered that ingression of primary mesenchyme cells occurs just as they replace one kind of cadherins by cell-matrix adhesion molecules.
Rachel Fink's discovery is an excellent example how changes in gene expression can cause cells to change locations inside the embryo. How do genes cause structure?

H. V. Wilson discovered that sponges can be separated into their individual cells, and then fairly quickly rearrange to re-form their anatomical patterns.
This shocked most embryologists, who have never really come to terms with it.
It implies that locations of differentiated cells in the body (even in our bodies)
result from active cell movements, not just from where the cells differentiated.
* Can you invent alternative theories to explain it? Or experiments to test them?
Holtfreter added five main contributions to this subject:
1) He found that by simply making their water slightly more acid, frog and salamander gastrula and neurula stages will fall apart into individual healthy cells.
2) He surgically dissected out various germ layers and early organs (from amphibian embryos), dissociated them, and randomly mixed them with dissociated cells from other germ layers.
3) He discovered that dissociated amphibian embryo cells quickly rearrange (sort out), getting back together with other cells of the same type, AND re-create normal anatomical positions of differentiated cells relative to each other.
4) Did some of these experiments with amphibians whose cells had very different colors, and kept those color differences through the sorting-out, which disproved the (widespread) idea that cells were re-differentiating rather than rearranging.
5) Proposed the explanatory concept of tissue affinity (Gewebaffinität), which people equated to selective adhesion, but Holtfreter also meant chemotaxis, or anything like that.
This stimulated many others to try to isolate & identify organ-specific glues Aaron Moscona was one of the best of these. He discovered that proteolytic enzymes can cause dissociation of all differentiated cells of bird and mammal embryos, and that they will sort out by differentiated cell type (liver with liver, heart with heart). He then tried to discover selective cell-cell adhesion proteins, like the cadherins.
J. P. Trinkaus then made two of the most important contributions to this field:
1) Radioactive labeling of DNA of dissociated and mixed cells proved for all time that cell sorting is re-positioning of cells that had kept their original differentiated cell type. He was one of the first to use radioactive labels (in this case, tritiated thymidine) combined with radioautography to trace cell fates.)
2) He made the first time lapse movies of dissociated cells during the process of sorting out, in some cases using highly colored cell types as a further proof that cells were rearranging by cell type, and not redifferentiating as a result of changed position.
Actually, he had wanted to find re-differentiation, & was a little disappointed.
A mark of a great scientist that he conclusive disproved his favorite theory!

Unfortunately, Trinkaus then concluded that sorting out by dissociated cells doesn't tell us anything about normal embryonic development.
One argument he made was that some of the pairs of cell types that he filmed sorting out don't ever normally come in contact with each other.
Another argument was that there are no kinds of animals whose development starts with a random mixture of differentiated cell types, which then spontaneously sort-out to form their anatomy.
Not even sponges and hydroids develop that way, although their cells sort out so well, and so constantly, and even without being dissociated that they might as well develop that way, and would be capable of it.
Instead, sponges start out with a non-random pattern of differentiated cells, which then sort-out to completely different geometrical arrangements, that they would have reached equally well starting from absolutely any initial geometric arrangement.
Sponge anatomy is totally the result of ongoing active cell and skeleton
rearrangements, just as if some scientist had randomly dissociated them.
Their cells sort out all the time, as a normal part of their life.
This was discovered by Bond and Harris, who didn't expect it.
Steinberg made at least 6 major research contributions to this subject.
1) He related cell sorting to the more general problem of explaining why anatomy gravitates toward certain preferred geometric arrangements.
2) He proposed a plausible "thermodynamic" hypothesis that attracted other researchers and stimulated lots of research, and much-needed theoretical concepts.
This thermodynamic "Differential Adhesion Hypothesis" is explained and dramatically illustrated in Scott Gilbert's textbook, on pages 70-74 of 7th edition.
3) He focused attention on the question of what sort of mechanism can cause cells to gravitate to the same end result (geometric arrangement) even when they can reach this end result by any of 2, or 3, or any number of different intermediate pathways.
His proposed explanation was that multiple alternative pathways to the same end result
means that the system is minimizing thermodynamic free energy.
He hypothesized that cells are maximizing their energy of adhesion to each other.
4) Based on his DAH theory, Steinberg made the testable prediction that (for example)
if dissociated cartilage cells will sort out into the interior of a mass of heart cells,
and if heart cells will sort out into the interior of masses of dissociated liver cells,
then cartilage cells will sort out internal to liver cells.
If C sorts internal to B, and B sorts internal to A, then for any combinations of 3 cell types, C will always sort out to an internal position relative to A cells.
This transitive relationship should be true for all combinations of differentiated cells.
Steinberg tested mixing dissociated cells in every possible combination of about 8 differentiated cell types, and in every one of the 45 (?) combinations, this transitivity of sorting-out position was always confirmed.
5) The DAH theory also predicted that if you centrifuge multicellular balls of cells, then the amount of flattening should be inversely proportional to how adhesive the cells are to each other. Thus, balls of liver cells should be flattened more than balls of heart cells, because the liver cells are (hypothesized to be) less adhesive. Steinberg and Herbert Phillips measured amounts of centrifugal flattening for spheres of dissociated and reaggregated cell types, and for every differentiated cell type, the amount of flattening produced by a given amount of centrifugal force was what the DAH theory predicted (cell types that sort out more interior, flatten less).
6) Steinberg did a sabbatical year in Takiechi's laboratory in Japan, and they did an experiment using certain cancer cells that had lost their genes for cadherin (the most important of the cell-cell adhesion proteins), and transfected different numbers of copies of a cadherin gene into these cells. They then dissociated the cells and mixed them together randomly, to see if they would sort out from each other, and they did. This proved that differences of numbers of adhesion proteins is sufficient to cause (otherwise identical) cells to sort out.
paper by Foty and Steinberg, 2004
Ramsey Foty, co-author on this paper
Harris made about 4 contributions to this general subject:
1) He and Bond discovered that cells of healthy, living, undisturbed sponges undergo constant active rearrangement, as if they were sorting out, even when you don't dissociate them at all.
2) He invented the flexible substratum method, first using gels with embedded marker particles, 8 years later developing the silicone rubber method, and discovered traction & large differences in strengths of traction between differentiated cell types
3) He showed that haptotaxis results from repeated tug of war events, rather than from cells maximizing their areas of contact and adhesion to substrata. The ventral areas of most cells don't have any adhesions at all, even to the most adhesive materials.
4) He published the major conceptual challenge to Steinberg's Differential
Adhesion Hypothesis DAH , proposing that Steinberg and Phillips'
quantitative measures of cell-cell adhesion were really measures of
contractility of cells at the aggregate surface, and that differences in
strengths of contraction, rather than strengths of cell-cell adhesion,
are the real cause of inner-outer positioning of aggregated cells.
He also explained that the predicted and carefully confirmed transitivity
inner-outer positioning of cell types is equally predicted by any theory
based on any quantitative variable. "Work of adhesion" is a quantitative
variable; so is strength of cortical (interfacial) contraction.

Another relevant fact is that differentiated cell types have different kinds of cell-cell adhesion proteins; not different amounts of a single kind of adhesion molecule, which was the foundation and motivation of the DAH.
I also argued that thermodynamic free energy can give misleading results in a system to which unknown amounts of energy are being added.
I proposed homeostasis as a better analogy to the stability and spontaneous rearrangements and shape changes. Homeostasis depends on negative feedback to produce stable (but adjustable) states of counterbalance of opposed forces.
Why couldn't these conceptual disagreements have been settled, with one or the other side convincing the others?
Sometimes they were settled amicably:
* Trinkaus experimentally disproved his own favorite theory, of cell redifferentiation.
** Carter quickly accepted the tug-of-war explanation of how cells behave on adhesion grids and gradients, in place of his theory that cells were pulled by a thermodynamic maximization of cell-solid adhesion.
Or...
*** Wilson himself never accepted that cells rearrange by cell type, and that he had discovered it. He believed they must instead be re-differentiating, and spent the rest of his life trying to prove that, using methods that were not able to disprove or prove either alternative.
**** Holtfreter poured contempt both on Steinberg's theory, & on my criticisms of Steinberg's theory, and my proposed alternatives.
Holtfreter wrote me a 34 page letter of criticism and derision, which contained the following cartoon and quasi-poetry.

Some more excerpts from Holtfreter's letter are appended at the end of this web page
Holtfreter believed that theorizing is an absurd waste of time,
and Trinkaus tended to agree. Nevertheless Trinkaus helped and
encouraged me, remained good friends with Steinberg and wrote
the following in his autobiography.
(Which I highly recommend that you consider buying and reading)



From EMBRYOLOGIST. My Eight Decades in Developmental Biology, by John Philip Trinkaus. J&S Publishing Company Inc., Alexandria, Virginia, 2003. ISBN 1888308-14-1
Physics could not have developed without the profound and careful theoretical reasoning of people like Newton, Clark-Maxwell, Einstein and Feynman. Developmental Biology has this same need for careful creation and analysis of difficult concepts. But it isn�t getting it. Instead, most of the best researchers have been intolerant of theories, which they were proud to leave to extremists, who they proudly made fun of, instead of following their reasoning carefully enough to find the fallacies. More recently, we have been getting out-and-out confidence tricksters. (Names supplied on request.)
What do you think? About the usefulness of theories?
About useful functions of theories? About distraction by theories?
About whether gastrulation, neurulation, somite formation, shaping of organs including the skeleton are caused by changes in cell-cell adhesion, changes in cell contraction,
or other properties that can be discovered and measured by dissociating cells?
More from Holtfreter:



Review Questions
1) The locomotion of body cells is more like that of which category of amoebae? [Dictyostelium]2) Which specific differentiated cell type are capable of crawling locomotion? [all or nearly all]
3) Describe the forces that propel crawling tissue cells?
4) Will body cells move preferentially onto (and into) more materials to which they are more adhesive?
5) Is this preferential adhesive movement onto more adhesive materials ("Haptotaxis") because
cells are physically pulled by the physical process of forming more or larger adhesions?
(Logically analogous to a magnet pulling on a piece of iron, and vice versa).
6) By what method were the locations and directions of this force discovered?
7) Can cell traction forces move non-living materials around in the body?
6) What happens to fluorescently-labeled collagen injected into developing embryos?
7) Does the collagen need to be from the same or different species than the embryo that it is injected into?
[No]
8) * Many cancer have consistent abnormalities of shape and behavior, which are visible in tissue culture.
Cancer cells' locomotion and shapes are about the same when cultured on relatively adhesive as on
relatively non-adhesive plastics or other materials: what explanations can you hypothesize?
9) * Try to invent hypotheses to explain the following facts: Most non-cancerous cells become shaped differently when crawling on less adhesive surfaces, as compared with how they behave on glass or other sticky surfaces. On less adhesive surfaces, the normal cells mimic shapes and movements of cancer cells.
10) ** Cancer can be induced by several kinds of solid materials, including some forms of asbestos,
if these solids become imbedded in your tissues. ** Hypothesize the mechanism.
(Other materials can be safely implanted in your body with no danger: **but why?)
11) * Which is more true:
a) That cells pull themselves by adhesions to external materials, or
b) That cells are pulled by formation of adhesions (that expansion of adhesions exerts a pulling force)?
14) The "Differential Adhesion Hypothesis" explains sorting out of differentiated cells from one another by assuming which of the two alternatives in the preceding question?
15) Does the author of our textbook seem to accept the Differential Adhesion Hypothesis as having been proven true?
16) Compare the opinions of Hans Holtfreter and J. P. Trinkaus about this ("DAH") theory.
Compare their opinions about complicated theories, in general, as applied to embryology.
(Theories are/aren't helpful? Theories shouldn't be too complicated? Nobody can find out which theory is true? It's better for scientists to concentrate on finding out what is true, not what theories predict?)
17) A researcher named Gershman did cell-sorting experiments in which he randomly mixed cancerous and (equivalent) non-cancerous cells, and discovered that they consistently sort out from each other,
with the normal cells bunching up in the interior, surrounded by layers of cancer cells.
In terms of the "DAH" what does this imply about differences between normal and cancer cells?
18) True or false: The anatomy of every kind of multicellular animal consists of some particular geometric arrangement of differentiated cells??
19) Which of the following explanations for these geometric arrangements of differentiated cells is more accurate?
#I) Cell differentiation is controlled, by chemical gradients or other signals, so that embryonic cells at each location differentiate into whichever cell type should be located at that position, and then they stay there?
# II) Among the special properties of each differentiated cell type is the ability to move actively, and to find their way to the correct functional position where their own particular cell type should be located?
[Usually, Embryology is discussed as if #I of these two alternatives is true. The textbook usually assumes this, even in the sections on regeneration of flatworms and other animals. But in the section on sorting out by differentiated cells, #2 is treated as a significant
contributor to the formation of body anatomy. In sponges, #2 is the ONLY cause of anatomy; and for other kinds of animals,
development occurs by mixtures of both these categories of mechanism. But most people think only #1 is important.]
Wayne Brodland at the University of Waterloo in Ontario has a web demonstration of cell sorting, together with his views on what explanatory mechanism is true. (Scroll down to the middle of the page, just below the legend to Fig. 7).
[Yes; This is called Haptotaxis and was discovered by Stephen Carter]
[No; It is analogous to the result of a tug-of-war between opposite sides of each cell,
in which less traction can be exerted on the sides of each cell that are on the least adhesive material]
[Putting tissue culture cells on elastic gels and sheets of rubber, and making videos
or tracking movements of marker particles or wrinkles.]
[Yes]
What are two examples of structural materials being moved around inside living animals?
[Spicules in sponges, and extracellular structural proteins like collagen in vertebrates & probably all phyla.]
[It gets rearranged and realigned to form tendons, part of blood vessel walls, etc]
[It assumes the truth of b,
because there can't be a "work of adhesion" or "energy of adhesion, unless the formation of adhesions exerts a pulling force, that expands areas of adhesions.]
[He seems to be completely convinced by the evidence that the DAH is the true cause of cell sorting, and also the cause of many normal embryological phenomena.]
[They both think it's a mistake to worry or think too much about any complicated or abstract theories,
and that scientists should concentrate on facts and experiments that are directly understandable]
That the cancer cells are less adhesive? That cancer cells' contractility is abnormally weak?
That cancer cells have less cadherin on their outer surfaces?
[According to the DAH, if cancer cells sort out to an exterior position relative to the closest equivalent kind of non-cancerous cells, then that means that the cancer cells are less adhesive than the normal cells. Having fewer molecules of cadherins on the outside surfaces of their plasma membranes is one possible reason that would cause cells to be less adhesive.]
[True. Plus also geometrical arrangements of extra-cellular proteins and skeleton.]
The relevant paper is Brodland, G.W., 2002, "The Differential Interfacial Tension Hypothesis
(DITH): A Comprehensive Theory for Self-Rearrangement of Embryonic Cells
and Tissues," ASME Journal of Biomechanical Engineering, Vol. 124, pp.
188-197.